Impact Factor ISSN: 1837-9664

 Global reach, higher impact
Global reach, higher impactJ Cancer 2023; 14(11):2075-2084. doi:10.7150/jca.86304 This issue Cite
Research Paper
Chemical screening identifies the anticancer properties of Polyporous tuberaster
Eun Seon Song1, Moon Kyoung So1, Hyon Jin Park1, Haneur Lee1, Yun Haeng Lee1, Myeong Uk Kuk1, Jiho Park1, Hyung Wook Kwon1,2, Jaehyuk Choi1, ![]() , Joon Tae Park1,2
, Joon Tae Park1,2 ![]()
1. Division of Life Sciences, College of Life Sciences and Bioengineering, Incheon National University, Incheon, Korea.
2. Convergence Research Center for Insect Vectors, Incheon National University, Incheon 22012, Korea.
Received 2023-5-18; Accepted 2023-7-3; Published 2023-7-9
Abstract

Most conventional anticancer drugs cause resistance to chemotherapy, which has emerged as one of the major obstacles to cancer treatment. In order to address this issue, efforts have been made to select new anticancer compounds from natural sources. The aim of this study is to identify novel anticancer compounds from mycelial culture extracts belonging to Polyporus tuberaster (P. tuberaster). Here, we found that mycelial culture extracts of P. tuberaster cultured in PDB medium (pt-PDB) effectively inhibited cancer cell growth. pt-PDB reduced the growth of cancer cells through apoptosis induction and S-phase arrest. The anticancer efficacy of pt-PDB was not to limited to one type of cancer. Furthermore, unlike traditional anticancer medications, pt-PDB did not increase the proportion of side population (SP) cells, which plays a key role in the development of chemoresistance. Taken together, we discovered a novel anticancer drug candidate that has anticancer properties without increasing the proportion of SP cells. This new drug candidate can be used for the treatment of cancer, especially chemoresistant malignancies, and will provide a breakthrough in the treatment of chemoresistant cancer.
Keywords: polyporus tuberaster, mycelium culture extract, chemical screening
Introduction
Mushrooms have been historically valued as a food source due to their low fat and high protein contents, as well as a functional food beneficial to human health [1]. Mushrooms produce primary metabolites and secondary metabolites [2]. Primary metabolites include nucleic acids and proteins, while secondary metabolites include polysaccharides, terpenoids, alkaloids, lectins, and metal chelators [3]. Secondary metabolites, in particular, have been found to possess potent biological activities, including antibacterial, antiviral, and anticancer properties [3]. Among the activities, anticancer effect has received much attention, and several clinical trials are being conducted to evaluate the efficacy of the mushroom-derived compounds [4]. For instance, Lingzhi mushroom (Ganoderma lingzhi) was demonstrated in a phase 2 clinical trial to help maintain the quality of life of cancer patients after chemotherapy [5]. Similarly, orally administered Ganoderma lucidum has been shown to be beneficial in the treatment of colorectal cancer patients by acting as an immunomodulator [6]. In another phase 1 clinical study, the button mushroom was found to reduce prostate-specific antigen levels in the patients with recurrent prostate cancer [7]. Taken together, the promising results from these clinical studies suggest that mushrooms with anticancer properties may hold great potential for the treatment of cancer patients either as a standalone therapy or in combination with conventional anticancer drugs.
The life cycle of mushrooms consists of two distinct stages: the long-lived mycelial stage and the short-lived fruiting body stage [8]. As fruiting bodies are cultured on a solid medium, large-scale cultivation requires large space and time. In contrast, mycelium is composed of tubular threads known as hyphae, and possesses a porous structure, rendering it suitable for liquid cultivation [9]. Recently submerged culture of mycelia has been introduced for mass cultivation of mycelia, thereby enabling the low-cost production of secondary metabolites on a large scale [9]. Maximizing the mass production of secondary metabolites through liquid culture of mycelia can be achieved by manipulating process factors and medium components [10]. Therefore, in order to mass-produce secondary metabolites at low cost, liquid culture using mycelia is essential.
Drug resistance is a major impediment to successful chemotherapy and continues to pose a challenge in cancer treatment [11]. Despite remarkable advances in conventional chemotherapy, the overall mortality rate of cancer patients remains high due to the emergence of drug-resistant cells [12]. Numerous studies are ongoing to identify novel mechanisms addressing drug resistance. Recent studies have revealed that side population (SP) cells play an important role in developing drug resistance and inducing relapse of all malignancies during chemotherapy [13, 14]. SP cells have multidrug resistance due to overexpression of the ATP-binding cassette (ABC) transporter, which plays a role in drug efflux [15, 16]. Given these results, finding drugs that do not increase the proportion of SP cells may reduce drug resistance, making it an effective chemotherapy option for cancer patients.
The genus Polyporus belong to the family Polyporaceae is a kind of wood rot mushroom [17]. Recently, Polyporous parvovarius was found to inhibit tumor cell proliferation and promote apoptosis [18]. However, investigations into the anticancer properties of other Polyporus species have not been extensively conducted despite the genus consisting of over 250 species. This study aimed to determine whether mycelial culture extracts of Polyporus tuberaster (P. tuberaster) possess anticancer properties. Our findings demonstrate that P. tuberaster mycelium culture extracts exhibit anticancer properties without inducing drug resistance.
Materials and Methods
Mycelium liquid culture
Polyporus tuberaster (Accession number: KMRB 18083112) used in this study was sourced from Korea Mushroom Resource Bank (KMRB, Seoul, Korea). Four media were used for mycelium culture (Table 1).
Names and components of the four media used in this study
| Media name | Components |
|---|---|
| PDB | • dextrose (215530; BD Difco, Franklin, NJ, USA) 20 g/l • potato starch (S2004; Sigma, Saint Louis, MO, USA) 4 g/l |
| MYB | • malt extract (218630; BD Difco) 20 g/l • yeast extract (212750; BD Difco) 2g/l |
| MEB | • maltose (216830; BD Difco) 1.8 g/l • dextrose (215530; BD Difco) 6 g/l • malt extract (218630; BD Difco) 6 g/l • yeast extract (212750; BD Difco) 2g/l |
| DYB | • dextrose (215530; BD Difco) 20 g/l • yeast extract (212750; BD Difco) 2g/l |
Extracts were produced from mycelium cultures as described in previous studies [19]. Briefly, precultured mycelial discs (5 mm) were inoculated into each Erlenmeyer flask with 700 ml of medium. The flasks were then incubated for 60 days at 25 °C while being shaken at 150 × g. To obtain a culture medium, a liquid culture medium containing mycelium was filtered through Miracloth (475855-1R; Calbiochem, La Jolla, CA, USA). The culture filtrate was separated and freeze-dried. Dried filtrates were soaked in 100% ethanol. After filtering the extract with 0.45 μm membrane filters (HAWP05000; Sigma) to remove the remaining residue, the ethanol was evaporated. The remaining extract was diluted in 500 mg/ml dimethyl sulfoxide (41639; Sigma).
Cell Culture
Seven different cell lines were used and detailed information is provided in Table 2. Cells was cultured as described previously [18].
Names and details of the 7 cell lines used in this study
| Cell line name | Catalogue number | Source |
|---|---|---|
| HeLa cells | A1100001; Thermo Fisher Scientific, Waltham, MA, USA | Cells were established from the tumor of a 31-year-old woman with a cervical carcinoma [20]. |
| BEAS-2B | CRL-9609™; American Type Culture Collection, Manassas, VA, USA | Cells were established from normal bronchial epithelium obtained from autopsy of non-cancerous individuals. Then, cells were infected with a replication-defective SV40/adenovirus 12 hybrid and cloned [21]. |
| NCI-H1299 | CRL-5803™; American Type Culture Collection | Cells were established from the tumor of a 43-year-old man with carcinoma [22]. |
| HCC15 | ACC 496; Deutsche Sammlung von Mikroorganismen und Zellkulturen, Braunschweig, German | Cells were established from the tumor of a 47-year-old man with non-small cell lung carcinoma (subtype squamous carcinoma) [23]. |
| A549 | CRL-185™; American Type Culture Collection | Cells were established from the lung tissue of a 58-year-old man with lung cancer [24]. |
| PC-9 | 90071810; European Collection of Authenticated Cell Cultures, Salisbury, UK | Cells were established from the tumor of a 45-year-old man with lung adenocarcinoma [25]. |
| NCI-H2009 | CRL-5911™; American Type Culture Collection | Cells were established from the tumor of a 68-year-old woman with stage 4 adenocarcinoma [26]. |
Cell proliferation assay
At 2,000 cells per well, HeLa cells were plated on 96-well plates. Extracts of P. tuberaster mycelium grown in four separate culture media (DYB, MEB, MYB and PDB) were diluted to a final concentration of 0.1 mg/ml. Four days after starting drug treatment, cell proliferation was assessed. A cell proliferation assay based on a DNA content was utilized to measure the number of cells [27]. Briefly, 50 μl of 0.2% SDS was added to each wells and incubated at 37 °C for 1 h. Then, 150 μl of SYBR Green I nucleic acid gel stain (1:1,000 in D.W., S-7567, Molecular Probes, Eugene, OR, USA) was added to each of the 96 wells. Fluorescence intensity was quantified to determine cell number using a VICTOR Multilabel Plate Reader (2030-0050; PerkinElmer, Waltham, MA, USA).
Determining pt-PDB's cytotoxicity
At 2,000 cells per well, HeLa cells were plated on 96-well plates. At dilutions of 0.1 to 0.5 mg/ml, pt-PDB was used. A cell proliferation assay based on a DNA content was utilized to measure the number of cells [27]. Four days after starting drug treatment, cell proliferation was assessed. Cell proliferation was measured at each dose and its values were normalized to those in DMSO in order to evaluate the viability of the cells.
Determining pt-PDB's optimal concentration to inhibit cell proliferation
At 2,000 cells per well, HeLa cells were plated on 96-well plates. Cell proliferation was assessed 1, 2, 3 and 4 days after treatment at concentrations of 0.02 to 0.1 mg/ml.
Trypan blue staining
Trypan blue staining with 0.4% trypan blue was assessed by a Cedex HiRes Analyzer (05650216001; Roche, Basel, Switzerland). Brightfield cell images were automatically captured on a Cedex HiRes analyzer.
Apoptosis assay
Apoptosis assay using the FITC Annexin V Apoptosis Detection Kit (556547; BD Biosciences, Franklin Lakes, NJ, USA) was performed as previously described [28].
Cell cycle assay
Cell cycle assay was performed as previously described [29]. In brief, cells were stained with 50 g/ml propidium iodide (PI, P4170-10MG; Sigma) after being fixed with 70% ethanol.
Western blot analysis
Western blot analysis was performed as described previously [18]. Primary antibodies used in this study p21Cip1 antibody (sc-6246; 1:500 dilution; Santa Cruz Biotechnology, Dallas, TX, USA), phospho-retinoblastoma (Rb) antibody (sc-271930; 1:500 dilution; Santa Cruz Biotechnology), phospo-ERK antibody (9101s; 1:500 dilution; Cell signaling technology, Danvers, MA, USA), p-MEK antibody (9121; 1:500 dilution; Cell signaling technology), Cleaved-Caspase 8 (9746s; 1:500 dilution; Cell signaling technology), Cleaved-Caspase 9 (9502S; 1:500 dilution; Cell signaling technology) and HRP-conjugated β-actin (sc47778; 1:1000 dilution; Santa Cruz Biotechnology). Secondary antibodies used in this study HRP conjugated anti-mouse antibody (sc-516102; 1:1000 dilution; Santa Cruz Biotechnology) and HRP conjugated anti-rabbit antibody (sc-2357; 1:1000 dilution; Santa Cruz Biotechnology).
Side population (SP) assay
5 μg/ml Hoechst 33342 (H3570; Thermo Scientific, Waltham, MA, USA) or 5 μM verapamil (V4629; Sigma) were applied to 1 × 106 cells for 90 min at 37 °C. Cells were stained with 1 μg/ml PI on ice for 5 min after being washed with PBS. LSR II (BD Biosciences) was used to analyze flow cytometry data.
Statistical analyses
The statistical analysis was carried out using a statistical software program (SigmaPlot 12.5; Systat Software, San Jose, CA, USA). Student's t-test was used to determine whether differences were significant.
Results
Chemical screening of P. tuberaster mycelium culture extracts for antitumor activity
A screening technique, a method of counting cells based on their DNA content, was used to find mycelial culture extracts of P. tuberaster that significantly slowed the growth of Hela cells [30]. Four extracts were produced from the mycelial culture of P. tuberaster, which was grown in four distinct media: DYB, MEB, MYB and PDB. Hela cells were treated with each extract at a dose of 0.1 mg/ml. The cell proliferation inhibitory effect was assessed on the 4th day of treatment. Paclitaxel, a drug commonly used to treat various types of tumors was used as a positive control [31]. The cell proliferation inhibition effects of mycelial culture extracts from MEB, MYB, and PDB media were significant when compared to the DMSO control (Fig. 1A). However, the mycelial culture extract in the DYB did not significantly inhibit cell proliferation. Mycelial culture extract in PDB medium (a.k.a., pt-PDB) was selected for further investigation because it reduced cell proliferation even more effectively than paclitaxel (positive control) (Fig. 1A).
Chemical screening of P. tuberaster mycelium culture extracts for antitumor activity. (A) Four extracts were produced from P. tuberaster mycelium grown in four distinct culture media (DYB, MEB, MYB and PDB). Hela cells were treated with each extract at a dose of 0.1 mg/ml. The cell proliferation inhibitory effect was evaluated on the 4th day of treatment. Paclitaxel was used as a positive control group. Student t-test, **P < 0.01, student t-test. Means ± S.D., n = 10. (B) Determining pt-PDB's cytotoxicity. Cell viability was assessed by measuring cell proliferation at 0.1-0.5 mg/ml pt-PDB concentrations and comparing the results to those in DMSO. Means ± S.D., n = 10. R2 (coefficient of determination) = 0.9241. (C) Determining pt-PDB's optimal concentration to inhibit cell proliferation. Cell proliferation was assessed 1, 2, 3 and 4 days after treatment at concentrations of 0.02 to 0.1 mg/ml. *P < 0.05, **P < 0.01, student t-test. Means ± S.D., n = 10.
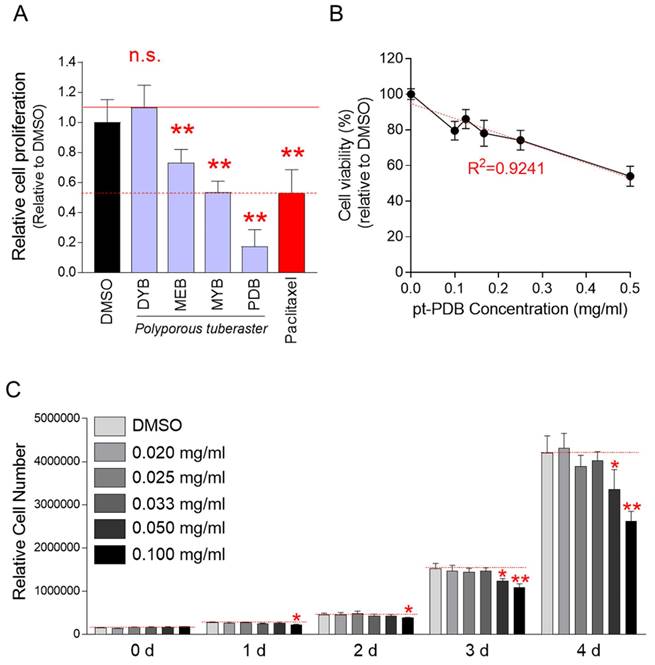
Although pt-PDB showed an inhibitory effect on cell growth, it was not rule out the possibility that this effect was due to cytotoxicity rather than an anticancer effect. To rule out this option, cell viability was assessed at doses of 0.1-0.5 mg/ml of pt-PDB. The decrease in cell viability was concentration dependent (Fig. 1B). R2 value, coefficient of determination, was 0.9241 indicating that the ability of pt-PDB to limit cell proliferation was not a result of cytotoxicity but rather an anticancer effect (Fig. 1B).
We then investigated the optimal pt-PDB concentration that could effectively reduce cell proliferation. Cell proliferation was assessed 0, 1, 2, 3 and 4 days after treatment at concentrations of 0.02 to 0.1 mg/ml. Compared to the DMSO control, pt-PDB concentrations below 0.05 mg/ml did not show a significant decrease in cell proliferation (Fig. 1C). However, a pt-PDB concentration of 0.05 mg/ml significantly decreased cell proliferation from day 3 after drug treatment (Fig. 1C). Moreover, the pt-PDB concentration of 0.1 mg/ml used for chemical screening showed a significant decrease in cell proliferation from day 1 after drug treatment (Fig. 1C). Since a pt-PDB concentration of 0.1 mg/ml inhibited cell proliferation from day 1 after drug treatment, 0.1 mg/ml was selected as the optimal pt-PDB concentration and applied to all subsequent experiments.
pt-PDB exerts anticancer activity through apoptosis induction and S-phase arrest
The anticancer activity of pt-PDB was determined by observing whether pt-PDB effectively kills cancer cells. Trypan blue staining was used to discriminate dead cells from live cells after pt-PDB treatment [32]. In the DMSO control, all cells were negative for trypan blue staining indicating live cells (Fig. 2; green squares represent live cells). However, in the pt-PDB group, cells positive for trypan blue staining were observed along with cells negative for trypan blue staining (Fig. 2; green squares represent live cells and red squares represent dead cells).
Determination of anticancer activity of pt-PDB by trypan blue staining. Cells were stained with 0.4% trypan blue and brightfield cell images were captured using a Cedex HiRes analyzer. Green squares represent live cells and red squares represent dead cells. Scale bar 10 μm.

Observation of dead cells by pt-PDB treatment led us to investigate the proportion of cells killed by apoptosis among dead cells, because apoptosis is a programmed cell death process, and effective elimination of cancer cells by apoptosis has been a critical goal of cancer treatment [33]. Apoptotic cells were discriminated from necrotic cells using annexin V/propidium iodide (PI) staining [34]. Compared to the DMSO control, pt-PDB treatment significantly increased the proportion of late apoptotic cells that were annexin V/PI-double positive (Fig. 3A). These results suggest that pt-PDB induces the release of pro-inflammatory cellular contents and membrane damage leading to late apoptotic cells.
pt-PDB causes apoptosis and S-phase arrest in order to inhibit cell proliferation. (A) Using annexin V and propidium iodide labeling, flow cytometric examination of apoptotic cells was carried out. Late apoptotic cell populations are represented by red squares. **P < 0.01, student's t-test. Means ± S.D., n = 3. (B) Effect of pt-PDB on the level of protein expression in the apoptotic pathway. Caspase 9 and Caspase 8 in their cleaved form. (C) Using PI labeling, flow cytometric examination of cell cycle was carried out. **P < 0.01, student's t-test. Means ± S.D., n = 3. (D) Effect of pt-PDB on the level of protein expression in the cell cycle pathway. p21Cip1 (p21), phospho-Rb, phospho-ERK, and phospho-MEK.
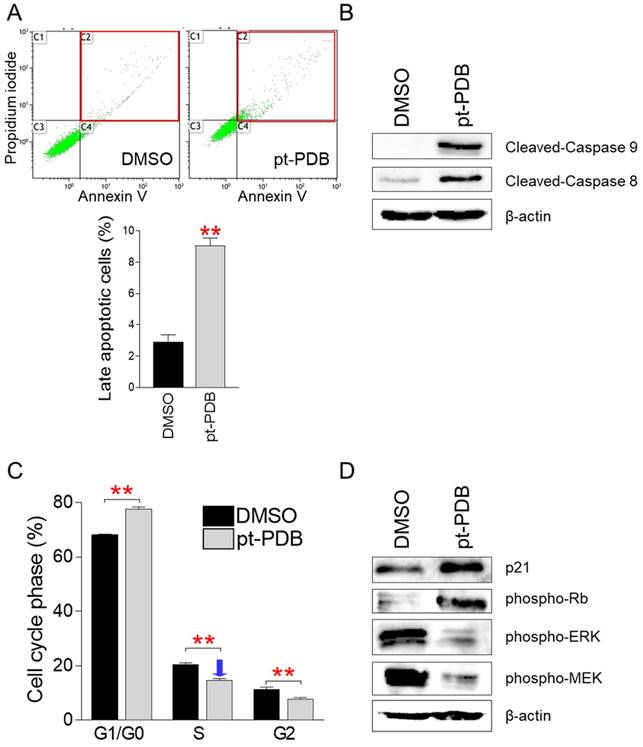
To further support the induction of apoptosis by pt-PDB, we examined the cleavage of caspase 9, which acts as an initiator of intrinsic apoptosis, and caspase 8, which acts as an initiator of death receptor-induced apoptosis [35]. Caspase 9 cleavage was much higher in pt-PDB compared to the DMSO control, indicating that the pt-PDB initiated intrinsic apoptosis (Fig. 3B). In addition, cleavage of caspase 8 was much higher in pt-PDB compared to DMSO group, indicating that pt-PDB executed death receptor-induced apoptosis (Fig. 3B).
Due to aberrant activation of cell cycle components, cancers exhibit deregulated cell proliferation [25]. Therefore, one of the key indicators for demonstrating the efficacy of anticancer effect is to determine whether a candidate anticancer agent induces cell cycle arrest, particularly S-phase arrest [26, 27]. We then investigated whether pt-PDB inhibits cell proliferation through triggering S-phase arrest. Compared to the DMSO control, cells treated with pt-PDB significantly increased the percentage of cells in G1/G0 phase from 68.1% to 77.6% (Fig. 3C). Moreover, pt-PDB significantly lowered the S-phase percentage from 20.5% to 14.6%, suggesting that pt-PDB inhibits cell proliferation by causing S-phase arrest (Fig. 3C).
The confirmation that pt-PDB inhibits cancer cell proliferation by inducing S-phase arrest led us to investigate how pt-PDB affects the expression levels of cell cycle regulatory proteins. p21Cip1 (p21) and phosphorylated form of retinoblastoma (p-Rb) are generally known proteins involved in S-phase arrest [36, 37]. Therefore, we examined the effect of pt-PDB treatment in the expression levels of p21 and p-Rb. Both p21 and p-Rb expressions were markedly increased by pt-PDB treatment, indicating that pt-PDB triggers S-phase arrest through activating p21-RB pathways (Fig. 3D). Extracellular signal-regulated kinase (ERK) and mitogen-activated protein kinase (MEK) are important controllers of S-phase entry signals during the cell cycle [38, 39]. Therefore, we investigated the expression levels of phospho-ERK (p-ERK), an active version of ERK, and phospho-MEK (p-MEK), an activated form of MEK. After pt-PDB treatment, the expression of p-ERK and p-MEK decreased, indicating pt-PDB treatment prevented the cell cycle progression (Fig. 3D).
pt-PDB prevents the proliferation of cancer cells derived from lung cancer
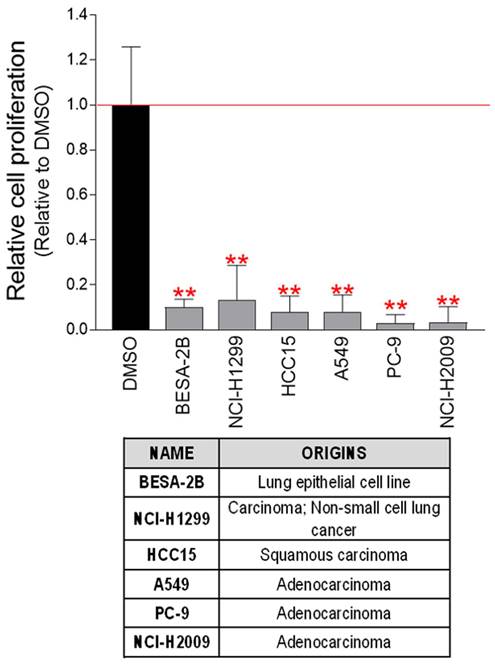
Validation of the anticancer effect of pt-PDB in cervical cancer-derived Hela cells led us to investigate whether the anticancer activity of pt-PDB extends to other cancer cell lines derived from surgically resected human tumor samples. Therefore, the effect of pt-PDB was investigated in six cancer cell lines derived from human lung cancer (BESA-2B, NCI-H1299, HCC15, A549, PC-9, NCI-H2009). Cancer cell growth was significantly reduced in all six lung cancer cell lines treated with pt-PDB (Fig. 4). These results suggest that the anticancer properties of pt-PDB can be used not only in cells derived from a single cancer species, but also in cell lines derived from other types of cancer.
pt-PDB has an anticancer effect on several lung cancer-derived cells. Six lung cancer-derived cells were exposed to pt-PDB diluted to a concentration of 0.1 mg/ml in the medium. The cell proliferation inhibitory effect was assessed on the 4th day of treatment. **P < 0.01, student t-test. Means ± S.D., n = 10.
pt-PDB does not increase the proportion of SP cells
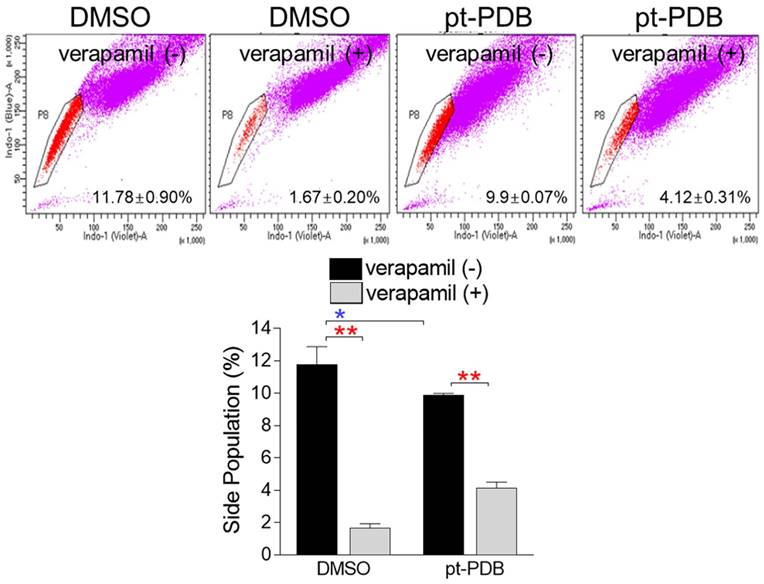
The emergence of malignant tumors resistant to chemotherapy is one of the major problems that make cancer treatment difficult [11]. SP cells, like chemoresistant cells, have the property of effluxing drugs via ABC transporters [40]. When stained with Hoechst 33342 dye, SP cells are distinguished from other cells because they rapidly efflux dye via ABC transporters [41]. However, SP cells lose this ability when exposed to ABC transporter inhibitors such as verapamil as they are unable to efflux the dye through the transporter [41]. Paclitaxel, the most popular anticancer drug, is very effective in treating various cancers, but like other chemotherapeutic agents, it also has a fatal disadvantage of inducing drug resistance by increasing the proportion of SP cells [39, 42]. We then looked at whether pt-PDB had side effects such as increasing the proportion of SP cell population. SP cells accounted for 11.78% of the total cell number in DMSO control group (Fig. 5). Verapamil significantly reduced the percentage of SP cells in DMSO control, indicating that the selected cells displayed SP cell characteristics (Fig. 5; red asterisk). Moreover, SP cells accounted for 9.9% of the total cell number in pt-PDB treated-group (Fig. 5). Verapamil significantly dropped the percentage of SP cells to 4.11% in pt-PDB group, proving that the selected cells had SP cell properties (Fig. 5; red asterisk). However, when comparing the ratio of SP cells between DMSO and pt-PDB groups, the percentage of SP cells in pt-PDB group was rather decreased compared to DMSO group (Fig. 5; blue asterisk). Taken together, our findings suggest that pt-PDB may be effective in treating chemoresistant cells because it exerts anticancer effects without increasing the proportion of SP cells.
pt-PDB does not increase the proportion of side population (SP) cells. Cells were stained using Hoechst 33342. Verapamil was used to prevent the efflux of Hoechst 33342. The side population was analyzed using flow cytometry. Side populations in the flow chart were placed using pentagon gates. *P < 0.05, **P < 0.01, student t-test. Means ± S.D., n = 3.
Discussion
Mushrooms produce secondary metabolites, which offer potential therapeutic benefits, including antioxidant and anticancer abilities [43]. Some secondary metabolites have been found to be relatively non-toxic and show potential for use as therapeutic drugs [44]. In this study, the mycelia of P. tuberaster were cultured using 4 distinct media (DYB, MEB, MYB, PDB). The culture extracts from DYB did not exhibit any anticancer activity, while the extracts from MEB, MYB, and PDB showed significant anticancer activities. This observation aligns with previous studies indicating that different supplements have been found to have a significant impact on mushroom yield and quality including secondary metabolites [45]. Notably, the culture extract of PDB showed the highest anticancer activity among all media tested. This can be attributed to the finding that the PDB medium contains only potato starch, whereas the DYB, MEB and MYB medium do not contain potato starch. Based on these results, it can be suggested that a medium containing potato starch creates an optimal medium condition for P. tuberaster to produce secondary metabolites with anticancer activity. This study is, to the best of our knowledge, the first to determine optimal media conditions for the formation of secondary metabolites with anticancer activity in P. tuberaster. If the composition of the PDB medium is further improved through processes such as spent medium analysis, it will a turning point in establishing medium conditions in which P. tuberaster or other mushrooms can generate secondary metabolites having anticancer activity.
Chemotherapy, which eliminates rapidly dividing cancer cells, is one of the most successful treatments for major cancers [46]. However, chemotherapy is successful in the early stages of cancer but loses effectiveness when cancer cells become chemoresistant [47]. For example, paclitaxel effectively kills cancer cells in the early stages of treatment, but loses its efficacy in the later stages [48]. Therefore, the discovery of drugs that do not cause drug resistance will open a new era in cancer treatment. Recent studies have demonstrated that SP cells are sources of drug resistance during chemotherapy [49, 50]. SP cells activate drug efflux channels such as ABC transporter to efflux various anticancer drugs. As a result, SP cells develop resistance to chemotherapy and eventually contribute to recurrent cancers [51, 52]. The novelty of this study lies in the fact that pt-PDB has anticancer properties, but unlike conventional anticancer drugs, it does not increase the proportion of SP cells. More research is needed to determine which active ingredient present in pt-PDB plays a key role in producing these effects, but the active ingredient alone or in combination with other anticancer drugs will be very effective in treating chemoresistant cancers.
Eukaryotic cells can kill themselves through the process of apoptosis without triggering an inflammatory response [53]. Since avoidance of apoptosis is an important characteristic of cancer cells, induction of apoptosis has been used as an effective method to prevent cell proliferation [33]. Another hallmark of cancer cells is dysregulation of cell cycle progression due to defects in regulatory proteins that drive cell cycle progression [54]. Therefore, controlling cell cycle progression has been considered a crucial objective of cancer treatment [54]. Here, pt-PDB induced apoptosis and S-phase arrest to kill cancer cells. These favorable results were not limited to a specific type of cancer cell and were effective for other cancer cell types as well. Based on the results that pt-PDB effectively controls the two most prominent characteristics of cancer cells, we suggest that pt-PDB should be first considered as an alternative treatment option for cancer treatment.
A new method called mycelial liquid culture allows mycelia to grow in liquid without the need for traditional agar plates [55, 56]. Compared to commonly used solid medium culture, mycelial liquid culture has the advantage of requiring less manufacturing time and culture space [56]. Mycelial liquid culture also allows rapid colonization of mycelia and scale-up of cell cultures, leading to low-cost mass production of secondary metabolites [57]. Therefore, the production of high value-added materials such as polysaccharides, pharmaceuticals, and enzymes is achieved through liquid culture of mycelia [56]. Here, liquid culture of P. tuberaster in PDB medium produced secondary metabolites with anticancer potential. However, since single liquid cultures were performed on a laboratory scale, the total amount of secondary metabolites obtained was limited. If a large-scale liquid culture production method is established by changing process parameters such as pH, temperature, dissolved oxygen, and agitation speed, the amount of secondary metabolites produced during single liquid cultures can be raised to a level suitable for commercial production.
In summary, P. tuberaster mycelium culture extracts effectively inhibited cancer cell growth by inducing apoptosis and S-phase arrest. The significance of its anticancer effect was reinforced by the finding that pt-PDB did not increase the proportion of SP cells. These novel anticancer candidates can be produced at low cost through mass culture and can be used clinically for the treatment of cancer, especially chemoresistant malignancies.
Acknowledgements
Funding
This research was supported by an Incheon National University research grant (2023-0015).
Author Contributions
ESS, JC and JTP conceived of and designed the experiments. ESS, MKS, HJP, HL, YHL, and MUK performed the experiments. ESS and HWK analyzed the data. ESS, JC and JTP wrote and edited the paper.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Cheung PCK. Mini-review on edible mushrooms as source of dietary fiber: Preparation and health benefits. Food Science and Human Wellness. 2013;2:162-6
2. Ziaja-Sołtys M, Kołodziej P, Stefaniuk D, Matuszewska A, Jaszek M, Bogucka-Kocka A. Low-Molecular-Weight Secondary Metabolites from Fungi: Cerrena unicolor as a New Proposal of an Effective Preparation against Rhabditis Nematodes. Molecules. 2022;27:1660
3. Chugh RM, Mittal P, Mp N, Arora T, Bhattacharya T, Chopra H. et al. Fungal Mushrooms: A Natural Compound With Therapeutic Applications. Front Pharmacol. 2022;13:925387
4. Wasser SP. Medicinal Mushrooms in Human Clinical Studies. Part I. Anticancer, Oncoimmunological, and Immunomodulatory Activities: A Review. International journal of medicinal mushrooms. 2017;19:279-317
5. Liu J, Mao JJ, Li SQ, Lin H. Preliminary Efficacy and Safety of Reishi & Privet Formula on Quality of Life Among Non-Small Cell Lung Cancer Patients Undergoing Chemotherapy: A Randomized Placebo-Controlled Trial. Integrative cancer therapies. 2020;19:1534735420944491
6. Chen X, Hu ZP, Yang XX, Huang M, Gao Y, Tang W. et al. Monitoring of immune responses to a herbal immuno-modulator in patients with advanced colorectal cancer. International immunopharmacology. 2006;6:499-508
7. Twardowski P, Kanaya N, Frankel P, Synold T, Ruel C, Pal SK. et al. A phase I trial of mushroom powder in patients with biochemically recurrent prostate cancer: Roles of cytokines and myeloid-derived suppressor cells for Agaricus bisporus-induced prostate-specific antigen responses. Cancer. 2015;121:2942-50
8. Kües U. Life History and Developmental Processes in the Basidiomycete <i>Coprinus cinereus</i>. Microbiology and Molecular Biology Reviews. 2000;64:316-53
9. Sugihara TF, Humfeld H. Submerged culture of the mycelium of various species of mushroom. Applied microbiology. 1954;2:170-2
10. Rathore H, Prasad S, Kapri M, Tiwari A, Sharma S. Medicinal importance of mushroom mycelium: Mechanisms and applications. Journal of Functional Foods. 2019;56:182-93
11. Housman G, Byler S, Heerboth S, Lapinska K, Longacre M, Snyder N. et al. Drug resistance in cancer: an overview. Cancers. 2014;6:1769-92
12. Miller KD, Nogueira L, Mariotto AB, Rowland JH, Yabroff KR, Alfano CM. et al. Cancer treatment and survivorship statistics, 2019. CA: a cancer journal for clinicians. 2019;69:363-85
13. Phi LTH, Sari IN, Yang Y-G, Lee S-H, Jun N, Kim KS. et al. Cancer Stem Cells (CSCs) in Drug Resistance and their Therapeutic Implications in Cancer Treatment. Stem Cells Int. 2018;2018:5416923 -
14. Akbar Samadani A, Keymoradzdeh A, Shams S, Soleymanpour A, Elham Norollahi S, Vahidi S. et al. Mechanisms of cancer stem cell therapy. Clinica Chimica Acta. 2020;510:581-92
15. Hirschmann-Jax C, Foster AE, Wulf GG, Nuchtern JG, Jax TW, Gobel U. et al. A distinct "side population" of cells with high drug efflux capacity in human tumor cells. Proceedings of the National Academy of Sciences of the United States of America. 2004;101:14228-33
16. Clarke RB, Spence K, Anderson E, Howell A, Okano H, Potten CS. A putative human breast stem cell population is enriched for steroid receptor-positive cells. Dev Biol. 2005;277:443-56
17. Cui B-K, Li H-J, Ji X, Zhou J-L, Song J, Si J. et al. Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China. Fungal Diversity. 2019;97:137-392
18. Lee YH, Kim M, Park HJ, Park JY, Song ES, Lee H. et al. Chemical screening identifies the anticancer properties of Polyporous parvovarius. Journal of Cancer. 2023;14:50-60
19. Kim M, Ahn C, Kim C. Major Antioxidant Compound of Polyporus Parvovarius Culture Filtrate. Natural Product Communications. 2022;17:1934578X211072336
20. Akiyama S. HeLa cell lines. Methods in enzymology. 1987;151:38-50
21. Han X, Na T, Wu T, Yuan BZ. Human lung epithelial BEAS-2B cells exhibit characteristics of mesenchymal stem cells. PLoS One. 2020;15:e0227174
22. Li J, Guo Q, Lei X, Zhang L, Su C, Liu Y. et al. Pristimerin induces apoptosis and inhibits proliferation, migration in H1299 Lung Cancer Cells. Journal of Cancer. 2020;11:6348-55
23. Virmani AK, Fong KM, Kodagoda D, McIntire D, Hung J, Tonk V. et al. Allelotyping demonstrates common and distinct patterns of chromosomal loss in human lung cancer types. Genes, Chromosomes and Cancer. 1998;21:308-19
24. Yu M, Qi B, Xiaoxiang W, Xu J, Liu X. Baicalein increases cisplatin sensitivity of A549 lung adenocarcinoma cells via PI3K/Akt/NF-κB pathway. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie. 2017;90:677-85
25. Qian Y, Dai H, Li H. Low-doses of aspirin promote the growth of human PC-9 lung cancer cells through activation of the MAPK family. Experimental and therapeutic medicine. 2021;22:1440
26. Dogan Turacli I, Candar T, Yuksel BE, Demirtas S. Role of metformin on base excision repair pathway in p53 wild-type H2009 and HepG2 cancer cells. Human & experimental toxicology. 2018;37:909-19
27. Silva LP, Lorenzi PL, Purwaha P, Yong V, Hawke DH, Weinstein JN. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical chemistry. 2013;85:9536-42
28. Hingorani R, Deng J, McIntyre C, Mittar D. Detection of apoptosis using the BD annexin V FITC assay on the BD FACSVerse™ system. BD Biosciences, San Jose. 2011:1-12
29. Pozarowski P, Darzynkiewicz Z. Analysis of cell cycle by flow cytometry. Methods in molecular biology (Clifton, NJ). 2004;281:301-11
30. Silva LP, Lorenzi PL, Purwaha P, Yong V, Hawke DH, Weinstein JN. Measurement of DNA Concentration as a Normalization Strategy for Metabolomic Data from Adherent Cell Lines. Analytical Chemistry. 2013;85:9536-42
31. Sharifi-Rad J, Quispe C, Patra JK, Singh YD, Panda MK, Das G. et al. Paclitaxel: Application in Modern Oncology and Nanomedicine-Based Cancer Therapy. Oxidative medicine and cellular longevity. 2021;2021:3687700
32. Crowley LC, Marfell BJ, Christensen ME, Waterhouse NJ. Measuring Cell Death by Trypan Blue Uptake and Light Microscopy. Cold Spring Harbor protocols. 2016. 2016
33. Pfeffer CM, Singh ATK. Apoptosis: A Target for Anticancer Therapy. International journal of molecular sciences. 2018;19:448
34. Rieger AM, Nelson KL, Konowalchuk JD, Barreda DR. Modified annexin V/propidium iodide apoptosis assay for accurate assessment of cell death. J Vis Exp. 2011 p. 2597
35. McIlwain DR, Berger T, Mak TW. Caspase functions in cell death and disease. Cold Spring Harb Perspect Biol. 2013;5:a008656
36. Zhu H, Zhang L, Wu S, Teraishi F, Davis JJ, Jacob D. et al. Induction of S-phase arrest and p21 overexpression by a small molecule 2[[3-(2,3-dichlorophenoxy)propyl] amino]ethanol in correlation with activation of ERK. Oncogene. 2004;23:4984-92
37. Knudsen KE, Booth D, Naderi S, Sever-Chroneos Z, Fribourg AF, Hunton IC. et al. RB-dependent S-phase response to DNA damage. Mol Cell Biol. 2000;20:7751-63
38. Wu D, Chen B, Parihar K, He L, Fan C, Zhang J. et al. ERK activity facilitates activation of the S-phase DNA damage checkpoint by modulating ATR function. Oncogene. 2006;25:1153-64
39. Rescan C, Coutant A, Talarmin H, Theret N, Glaise D, Guguen-Guillouzo C. et al. Mechanism in the sequential control of cell morphology and S phase entry by epidermal growth factor involves distinct MEK/ERK activations. Mol Biol Cell. 2001;12:725-38
40. Murase M, Kano M, Tsukahara T, Takahashi A, Torigoe T, Kawaguchi S. et al. Side population cells have the characteristics of cancer stem-like cells/cancer-initiating cells in bone sarcomas. British journal of cancer. 2009;101:1425-32
41. Murota Y, Tabu K, Taga T. Requirement of ABC transporter inhibition and Hoechst 33342 dye deprivation for the assessment of side population-defined C6 glioma stem cell metabolism using fluorescent probes. BMC cancer. 2016;16:847
42. Shen Y, Yang L, Li L. Cancer stem-like cells contribute to paclitaxel resistance in esophageal squamous cell carcinoma. International journal of clinical and experimental pathology. 2022;15:183-90
43. Bhambri A, Srivastava M, Mahale VG, Mahale S, Karn SK. Mushrooms as Potential Sources of Active Metabolites and Medicines. Front Microbiol. 2022;13:837266
44. Bains A, Chawla P, Kaur S, Najda A, Fogarasi M, Fogarasi S. Bioactives from Mushroom: Health Attributes and Food Industry Applications. Materials. 2021;14:7640
45. Carrasco J, Zied DC, Pardo JE, Preston GM, Pardo-Giménez A. Supplementation in mushroom crops and its impact on yield and quality. AMB Express. 2018;8:146
46. Schirrmacher V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment (Review). International journal of oncology. 2019;54:407-19
47. Park JT, Chen X, Tropè CG, Davidson B, Shih Ie M, Wang TL. Notch3 overexpression is related to the recurrence of ovarian cancer and confers resistance to carboplatin. Am J Pathol. 2010;177:1087-94
48. Chun R, Garrett LD, Vail DM. Chapter 11 - Cancer Chemotherapy. In: Withrow SJ, Vail DM, editors. Withrow & MacEwen's Small Animal Clinical Oncology (Fourth Edition). Saint Louis: W.B. Saunders. 2007 p. 163-92
49. Visvader JE, Lindeman GJ. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nature reviews cancer. 2008;8:755-68
50. Alison MR, Lim SM, Nicholson LJ. Cancer stem cells: problems for therapy? The Journal of pathology. 2011;223:148-62
51. Goodell MA, Brose K, Paradis G, Conner AS, Mulligan RC. Isolation and functional properties of murine hematopoietic stem cells that are replicating in vivo. The Journal of experimental medicine. 1996;183:1797-806
52. Pattabiraman DR, Weinberg RA. Tackling the cancer stem cells - what challenges do they pose? Nature reviews Drug discovery. 2014;13:497-512
53. D'Arcy MS. Cell death: a review of the major forms of apoptosis, necrosis and autophagy. Cell biology international. 2019;43:582-92
54. Otto T, Sicinski P. Cell cycle proteins as promising targets in cancer therapy. Nature reviews Cancer. 2017;17:93-115
55. Wang S, Wang C, Cao H, Cui X, Guo H, Zheng W. et al. Comparing the Cosmetic Effects of Liquid-Fermented Culture of Some Medicinal Mushrooms Including Antioxidant, Moisturizing, and Whitening Activities. International journal of medicinal mushrooms. 2020;22:693-703
56. Bakratsas G, Polydera A, Katapodis P, Stamatis H. Recent trends in submerged cultivation of mushrooms and their application as a source of nutraceuticals and food additives. Future Foods. 2021;4:100086
57. Manan S, Ullah MW, Ul-Islam M, Atta OM, Yang G. Synthesis and applications of fungal mycelium-based advanced functional materials. Journal of Bioresources and Bioproducts. 2021;6:1-10
Author contact
![]() Corresponding authors: Jaehyuk Choi, Ph.D., Division of Life Sciences, College of Life Sciences and Bioengineering, Incheon National University, Incheon 22012, Republic of Korea. Tel: +82-32-835-8242; E-mail address: jaehyukcac.kr. Joon Tae Park, Ph.D., Division of Life Sciences, College of Life Sciences and Bioengineering, Incheon National University, Incheon 22012, Republic of Korea. Tel: +82-32-835-8841; E-mail address: joontae.parkac.kr.
Corresponding authors: Jaehyuk Choi, Ph.D., Division of Life Sciences, College of Life Sciences and Bioengineering, Incheon National University, Incheon 22012, Republic of Korea. Tel: +82-32-835-8242; E-mail address: jaehyukcac.kr. Joon Tae Park, Ph.D., Division of Life Sciences, College of Life Sciences and Bioengineering, Incheon National University, Incheon 22012, Republic of Korea. Tel: +82-32-835-8841; E-mail address: joontae.parkac.kr.