Impact Factor ISSN: 1837-9664

 Global reach, higher impact
Global reach, higher impactJ Cancer 2017; 8(13):2424-2435. doi:10.7150/jca.19639 This issue Cite
Research Paper
Loss of Tff1 Promotes Pro-Inflammatory Phenotype with Increase in the Levels of RORγt+ T Lymphocytes and Il-17 in Mouse Gastric Neoplasia
Mohammed Soutto1, 2*, Mohamed Saleh3, 6*, Mohamed S Arredouani4, Blanca Piazuelo5, Abbes Belkhiri2, Wael El-Rifai1, 2 ![]()
1. Department of Veterans Affairs, Tennessee Valley Healthcare System, Nashville, TN 37232;
2. Department of Surgery, Vanderbilt University Medical Center, Nashville, TN 37232;
3. Divison of Clinical Pharmacology, Vanderbilt University Medical Center, Nashville, TN 37232;
4. Division of Urology, Department of Surgery, Beth Israel Deaconess Medical Center and Harvard Medical School, Boston, MA 02115;
5. Division of Gastroenterology, Hepatology, & Nutrition, Vanderbilt University Medical Center, Nashville, TN 37232;
6. Department of Pharmacology and Toxicology, Faculty of Pharmacy, Mansoura University, Mansoura, Egypt 35516.
* equal contribution
Received 2017-2-13; Accepted 2017-5-18; Published 2017-7-22
Abstract
Background: TFF1 deficiency induces a mucosal pro-inflammatory phenotype that contributes to gastric tumorigenesis in mouse and human. Methods: We utilized the Tff1-KO mouse model to assess the impact of TFF1 loss on immune cells infiltration in the stomach. We used single cell suspension, flow cytometry, immunohistochemistry, and quantitative PCR (qPCR) assays. Results: The Tff1-KO gastric mucosa demonstrated high chronic inflammatory scores (score: 3-4) at age 2 months, which exacerbated at age 8 months (score: 4-6). We next used single-cell suspensions for flow cytometry analysis of total leukocytes (CD45+ cells), total T lymphocytes (CD45+CD3+cells), T cell subsets (CD4+, CD8+, and CD3+CD4-CD8-cells), and monocytes/macrophages (CD45+F4/80+cells). The results demonstrated an age-dependent (2 → 8 month age) significant increase of leukocytes (p<0.05), T cells (p<0.05), and monocytes/macrophages (p<0.001) in the gastric mucosa of the Tff1-KO mice, as compared to Tff1-WT. A similar increase was observed in blood samples (p<0.05). Using ionomycin to activate CD4+ splenocytes, the results indicated that Tff1-KO CD4+ splenocytes secreted higher levels of IL-17A (p<0.05 at 2 and p<0.001 at 8 months) and IL-17F (p<0.05 at 2 and 8 months) than Tff1-WT splenocytes. Conversely, Tff1-KO CD8+-cells secreted less IL-17F, but comparable levels of IL-17A. In addition, we detected a significant upregulation of Il-17 mRNA expression in gastric tissues in the Tff1-KO, as compared to Tff1-WT (p<0.001). Conclusions: The results identify TFF1 loss as a major pro-inflammatory step that modulates the tumor microenvironment and immune cell infiltration in the stomach. Furthermore, the data suggest that the increase of IL-17A and IL-17F in Th17 cells, derived from CD4+ T cells, reflects the chronic inflammation in gastric mucosa, whereas the absence of change of IL-17A and decrease of IL-17F in CD8+Tc17 cells suggest loss of cytotoxic function of CD8+Tc17 cells during gastric tumorigenesis of the Tff1-KO mice.
Keywords: TFF1, Immune cells, CD8, lymphocytes, Il-17, gastric cancer.
Introduction
The concept that interaction between immune cells and the microenvironment is actively involved in tumor development has gained extensive interest in recent years. The immune system is in fact an important determinant of the tumor microenvironment [1, 2]. Immune cells have the capability to eradicate malignancy through inflammation as a protective physiological response to repair damaged tissues and restore homeostasis. Alternatively, they can promote carcinogenesis through deleterious chronic inflammation that leads to accumulation of DNA damage and mutations [3, 4]. Chronic inflammation is characterized by a sequence of events produced in response to recognition of pathogens or tissue damage involving infiltration of immune cells and soluble mediators such as cytokines of the innate and adaptive immune system [5]. The presence and significance of leukocytic infiltration in developing neoplasms are now undisputed [6-8]. Certain leukocytes, cytotoxic T lymphocytes and natural killer cells, have the potential to eradicate malignant cells [9-11]. On the other hand, some leukocytic cell types, most notably innate immune cells such as immature myeloid cells, T cells, and macrophages, can instead potentiate tumor progression [9-11].
IL-17 is a family of cytokines composed of 6 members (IL-17A-F) [12]. While IL-17A and IL-17F are the signature cytokines defining CD4+ Th17 cells [13], it should be noted that IL-17 can also be produced by CD8+ Tc17 [14]. Previous studies have shown that Th17 cells and associated cytokines were present at significant levels during the progression and metastasis of gastric cancer [15, 16]. However, the role of Tc17 in tumor immunity against gastric cancer remains unclear. Of note, IL-17 (Tc17) promoted antitumor immunity against B16 melanoma in mice [17]. Th17 and Tc17 both express the transcription factor retinoic acid receptor-related orphan receptor gamma t (RORγt), which acts as a molecular determinant for their polarization and is necessary and sufficient for IL-17 expression [18, 19].
TFF1 is a small-secreted protein predominantly expressed by the gastric epithelia, in the upper portion of the glandular pits [20]. A number of studies have shown that TFF1 functions as a gastric tumor suppressor gene [21-26]. In human, more than half of gastric adenocarcinomas demonstrate downregulation of TFF1 expression by molecular mechanisms that include deletions, mutations, loss of heterozygosity, and promoter hypermethylation [27-30]. In a mouse model, loss of Tff1 in the Tff1-knockout (KO) leads to chronic inflammation and the development of gastric neoplasms at the antropyloric gastric regions [23, 24, 31]. The use of non-steroidal anti-inflammatory cox-2 inhibitor (celecoxib) in this mouse model led to a significant decrease in expression of inflammatory cytokines with 50% reduction in occurrence of gastric neoplasms [23]. However, the roles TFF1 in regulating inflammation and the development of cancer are not fully understood.
In this report, using the Tff1-KO mouse model, we characterized the immune cells in the gastric tissue microenvironment to identify cell populations that are involved in chronic inflammation and the development of gastric neoplasms in this model. Our findings demonstrate an increase in the number of infiltrating immune cells and secretion of IL-17 through Th17 immune cells in the Tff1-KO mice gastric tissues.
Materials & Methods
Animals
Animals and histologic evaluation
The Tff1-knockout (KO) and normal Tff1 wild-type mice (WT) [21] were used in this study. All animal studies were approved by the Institutional Animal Care and Use Committee at Vanderbilt University Medical Center. Following euthanasia, animals were dissected through midline incision of the abdomen. Stomachs from Tff1-KO and Tff1-WT were removed, cut along the lesser curvature, washed with PBS, and opened to lie flat. The stomachs were examined visually for abnormalities, size and number of individual gastric tumors and photographed. The stomach was cut into symmetrical halves where one-half was snap frozen and the other half was submerged in 10% buffered formalin solution, embedded in paraffin, and processed for standard H&E staining. Histopathology and chronic inflammation scores were evaluated at 2 months of age in Tff1-WT (n=29) and Tff1-KO (n=17) and at ≥8 months age in Tff1-WT (n=32) and Tff1-KO (n=122).
Immunohistochemical assessment
Formalin-fixed, paraffin-embedded sections of gastric tissues from Tff1-WT and Tff1-KO mice at age 2 and 8 months were cut at 5 microns. For CD3 staining, antigen retrieval was performed in pH 6.0- citrate buffer, by using a pressure cooker at 104°C for 20 minutes with a 10-minute bench cool down, followed by quenching with 0.03% H2O2 w/sodium azide for 5 minutes. After blocking with Dako X0909 serum-free protein block for 15 minutes, the samples were incubated with CD3 Dako cytomation rabbit polyclonal primary antibody at 1:100 dilution for an hour. For F4/80 staining, antigen retrieval was performed by incubating samples with Proteinase K for 5 minutes, and quenching with 0.03% H2O2 w/sodium azide for 5 minutes. Sections were incubated for one hour with rat anti-mouse F4/80 primary antibody at 1:100 dilution followed by a second incubation with rabbit anti-rat immunoglobulin at 1:100 dilution for 20 minutes. For detection of CD3 and F4/80, we used DAKO EnVision + HRP Labeled Polymer (Agilent Technologies, Santa Clara, CA) for 30 minutes followed by incubation with chromogen DAB+ for 5 minutes.
Flow cytometry of gastric and blood leukocytes
Enzymatic dissociation of gastric cells was performed on freshly resected antropyloric gastric tissues using 1 mg/mL collagenase A, 1 mg/ml collagenase B, and 100 µg/ml DNase I in RPMI 1640 medium with 5% FBS for 30 minutes at 37°C, with intermittent agitation. Homogenates were then filtered through a 70 µm cell strainer. Single cell suspensions were washed with cold PBS then stained for flow cytometry after blocking of Fc receptors with CD16/CD32 for 20 minutes at 4°C (BD Biosciences, clone 2.4G2). Antibodies used for flow cytometry were as follows: Brilliant Violet 510 (BV510)-conjugated anti-CD45 antibody (BioLegend, San Diego, CA, clone 30-F11), peridinin chlorophyll protein-cyanin-5.5 (PerCP-Cy5.5)-conjugated anti-CD3 antibody (BioLegend, clone 17A2), phycoerythrin-cyanin-7 (PE-Cy7)-conjugated anti-CD8 antibody (BioLegend, clone 53-6.7), allophycocyanin-Hilite-7 (APC-H7)-conjugated anti-CD4 antibody (BD Biosciences, clone GK1.5), Alexa Fluor 488-conjugated anti-F4/80 antibody (BioLegend, clone BM8). For live/dead cell staining, 7-aminoactinomycin-D (7-AAD) (eBioscience, San Diego, CA) was added to the cells just prior to analysis on a BD FACS Canto II™ system. Analysis was performed using BD FACSDiva software (BD Biosciences, Bedford, MA). Results were expressed as absolute number of cells in a specific population. Gastric tissue was consistent in weight among all groups. Gating was applied using fluorescence minus one (FMO) controls. The absolute number of infiltrating cells was calculated by multiplying each population cell number by the total number of live cells (assessed using trypan blue exclusion on a hemocytometer before staining) then dividing by the total number of live singlet cell populations obtained from flow cytometry analysis. In addition, we performed other methods to confirm our absolute cell counting by adding a known quantity (50 µl) of calibration (counting) beads to a known sample volume (CountBright™ Absolute Counting Bead, Life Technologies, Carlsbad, CA). For total blood staining, 100 µl of fresh heparinized blood was directly stained with 1.5 µl of each of the previously mentioned antibodies for 30 minutes at 4°C followed by washing with FACS buffer and then red cell lysis buffer was applied. Finally, 7-AAD was added to the cell pellet just prior to analysis on a BD FACS Canto II™ system.
Measurement of spleen IL-17A and IL-17F and detection of RORγt in blood and gastric tissues
Spleens were ground and filtered through a 70 μm cell strainer. RBCs were lysed using RBC lysis buffer (eBioscience). Isolation of renal leukocytes was processed as discussed in the previous step. Approximately 1x106 of either spleen of kidney cells were resuspended in RPMI medium supplemented with 5% FBS and stimulated with 2 µl of BD Leukocyte Activation Cocktail (ionomycin and phorbol 12-myristate 13-acetate (PMA) along with the Golgi inhibitor, brefeldin A) at 37°C for 5 hrs. Cells were washed and stained first with LIVE/DEAD® Fixable Violet dead cell stain (Invitrogen, Carlsbad, CA). The following surface antibodies were then added for 30 minutes: BV510-conjugated anti-CD45 antibody (clone 30-F11), PerCP-Cy5.5-conjugated anti-CD3 antibody (BioLegend, clone 17A2), and APC-H7-conjugated anti-CD4 antibody (BD Biosciences, clone GK1.5). Intracellular staining was then performed with the BD Cytofix/CytopermTM Plus fixation/permeabilization solution kit following the manufacturer's instructions (BD Biosciences) and using phycoerythrin (PE)-conjugated anti-IL-17A antibody (eBioscience, clone eBio17B7). We followed exactly the same protocol to detect RORγt in blood and gastric tissues, however, we did not stimulate with ionomycin/PMA. PE-conjugated anti-RORγt antibody (eBioscience, clone B2D) was used.
Quantitative real-time RT-PCR
Total RNA was isolated using the RNeasy Mini kit (Qiagen, Germantown, MD), and single-stranded cDNA was subsequently synthesized using the Advantage RT-for-PCR Kit (Clontech, Mountain View, CA). Genes specific for mouse primers were designed using the online software Primer 3 (http://frodo.wi.mit.edu/primer3/). The forward and reverse primers were designed to span two different exons for IL-17 gene; forward 5' CAGGACGCGCAAACATGA 3', reverse 5' GCAACAGCATCAGAGACACAGAT 3'. The primers were purchased from Integrated DNA Technologies (Coralville, IA). The qRT-PCR was performed using an iCycler (Bio-Rad, Hercules, CA), with the threshold cycle number determined by using a Bio-Rad CFX Connect Real-time System, with the threshold cycle number determined by Bio-Rad CFX manager software version 3.0. The results of the genes expression were normalized to housekeeping gene actin, as described previously. Expression ratios were calculated according to the formula 2(Rt-Et)/2(Rn-En) [32], where Rt is the threshold cycle number for the reference gene observed in the test samples, Et is the threshold cycle number for the experimental gene observed in the test samples, Rn is the threshold cycle number for the reference gene observed in the reference samples, and En is the threshold cycle for the experimental gene observed in the reference samples. Rn and En values were calculated as an average of all reference samples.
Statistical analysis
Using GraphPad Prism software (GraphPad Software, Inc., La Jolla, CA), a 2-tailed Student's t-test was used to compare the statistical difference between 2 groups. The differences were considered statistically significant when p value was <0.05.
Results
Loss of Tff1 expression induces infiltration of immune cells, chronic inflammation and tumorigenesis in mice
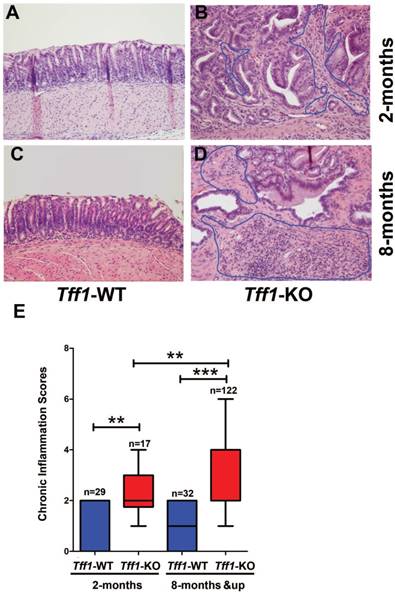
We have previously reported that gastric precancerous hyperplasia and low grade dysplasia lesions start at the age of 2 months and the establishment of high grade dysplasia and invasive adenocarcinoma occur at ≥ 6 months of age in Tff1-KO mice. In order to understand the role of immune cells in the gastric tumorigenesis cascade in this mouse model, we investigated the tissue microenvironment in the antra-pyloric area of the stomach in 2 and 8 month old Tff1-KO and Tff1-WT mice. By examining the gastric histopathology using H&E staining, we found limited or absent infiltration of immune cells in the antrum of Tff1-WT mice at all ages (Figure 1A&1C). Conversely, there was a substantial immune cell infiltration in low grade dysplasia of the antrum at the age of 2 months (Figure 1B) and high grade dysplasia at the age of 8 months (Figure 1D) in Tff1-KO mice. We next analyzed the chronic inflammatory scores in the gastric mucosa of the Tff1-KO and Tff1-WT mice of both ages. Our data indicated a significant increase of chronic inflammatory scores in 2-month-old Tff1-KO mice (p<0.01); this increase was more significant in 8-month-old Tff1-KO (p<0.001), as compared to Tff1-WT mice of matching age (Figure 1E). Notably, the inflammatory scores were low and remained unchanged in the gastric mucosa of the Tff1-WT mice at all ages (Figure 1E). These data strongly suggest that the inflammation observed in Tff1-KO mice antra-pyloric gastric tissues implicates infiltrating immune cells.
Loss of Tff1 promotes immune cells infiltration and chronic inflammation in premalignant gastric lesions. (A-D) H&E staining of representative histological features of gastric mucosa from wild-type and Tff1-knockout mice (2 and 8 months of age) showing infiltration of immune cells in Tff1-KO (B&D) but not in Tff1-WT (A&C). Original magnification, ×20. (E) Comparison of chronic inflammation in gastric tissues of matched ages (2 and 8 months) between wild-type and Tff1-knockout mice; each data point represents a single mouse, and the horizontal bars denote the mean value. **, p<0.01; ***, p<0.001.
Loss of Tff1 expression promotes infiltration of T lymphocytes and monocytes in gastric tissues
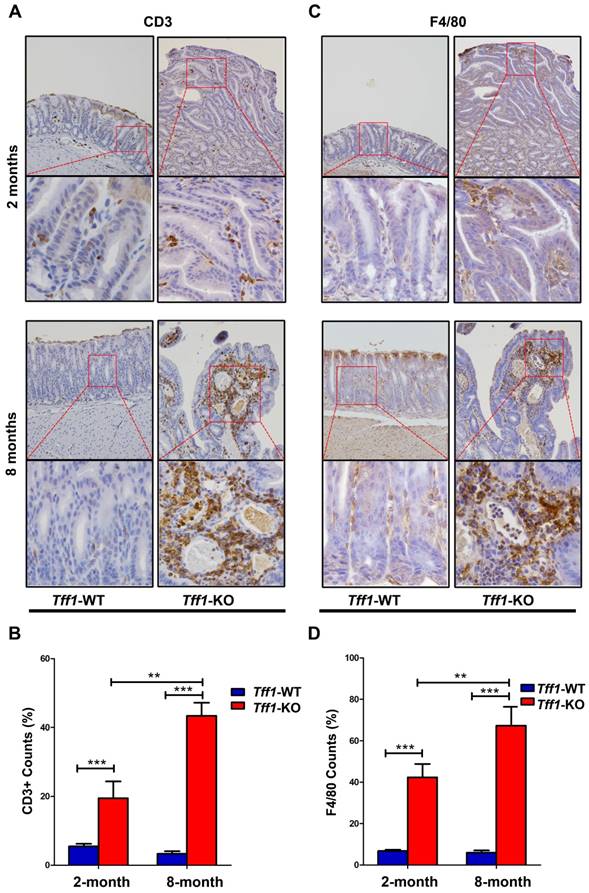
Several studies have shown that infiltration of immune cells can promote tumorigenesis in inflammation-associated cancers [33-35]. We, therefore, investigated if loss of Tff1 expression in mouse gastric tissue would exhibit increased immune cells infiltration and inflammation. We employed immunohistochemistry for lymphocytes and macrophages using CD3 and F4/80 antibody markers, respectively, in gastric tissues sections from 2 and 8 month old Tff1-WT and Tff1-KO mice. Our data indicated a significant presence of CD3+ T cells (Figure 2A&2B, p<0.001) and F4/80 macrophages (Figure 2C&2D, p<0.001) infiltrating in gastric mucosa of Tff1-KO more than in Tff1-WT mice. Of note, there were significantly more infiltrating CD3+ T lymphocytes and F4/80 macrophages in 8-month-old than in 2-month-old Tff1-KO mice (Figure 2B&2D, p<0.01). This increase of infiltrating immune cells was also localized more in the dysplastic than in non-dysplastic lesions in Tff1-KO gastric mucosa (Figure 2A&2C). These data strongly suggest that gastric tumorigenesis, due to loss of Tff1 expression, is associated with immune cells infiltration and inflammation in the gastric mucosa.
Immunohistochemical analysis for T lymphocytes and monocytes/macrophages in Tff1-WT and Tff1-KO gastric tissue. (A&C) Representative immunohistochemistry staining for the T cell marker CD3 (A) and F4/80 marker for macrophages (C) in gastric tissue of 2 and 8 month old mice (n=3 to 5 mice per group). (B&D) quantification analysis of CD3+ (B) and F4/80 (D) in four different random microscopic fields. The number of cells positive for CD3 and F4/80 was significantly higher in dysplastic lesion glands in Tff1-KO mice than normal glands in Tff1-WT (p<0.001). Original magnification at ×20 (top) and x40 (bottom).
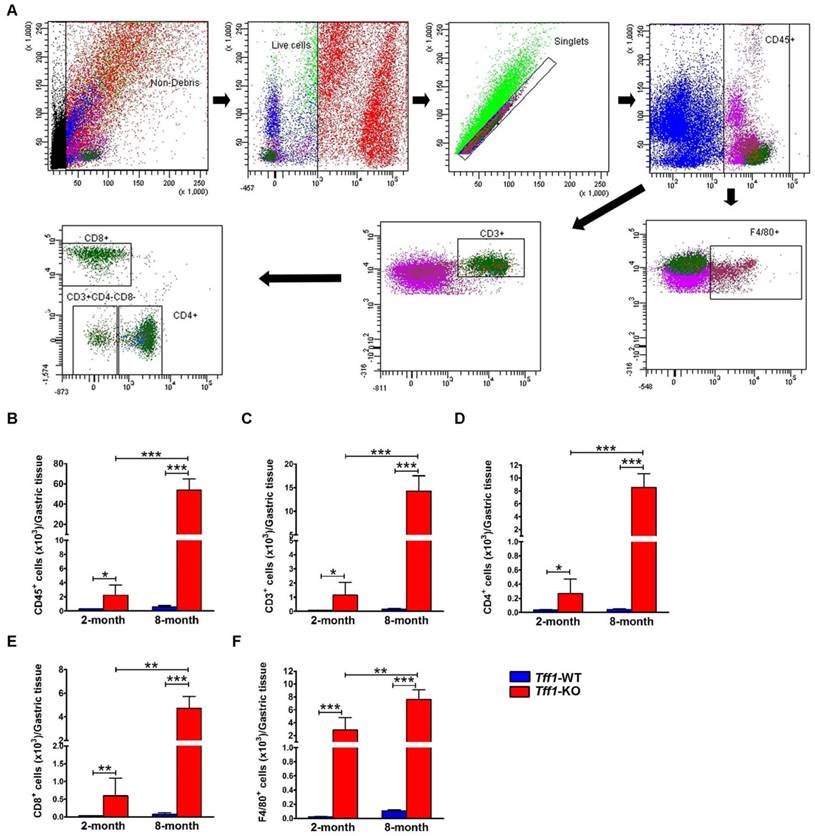
To validate the immunohistochemistry data, we evaluated the distribution of macrophages and T lymphocytes subpopulations. Using single-cell suspensions from gastric tissues of Tff1-WT and Tff1-KO mice, we performed flow cytometry to quantify total leukocytes (CD45+ cells), total T lymphocytes (CD45+CD3+ cells), T cell subsets (CD4+ and CD8+ cells), and monocytes/macrophages (CD45+F4/80+ cells) (Figure 3A). Consistent with the histological analysis and immunohistochemistry results, the flow cytometry plots and quantitative data showed a significant increase in total leukocyte infiltration in the stomach of 2 (p<0.05) and 8 (p<0.001) month old Tff1-KO mice as compared to Tff1-WT mice of matching age (Figure 3B). The flow cytometry data also indicated a significant increase of total T lymphocytes, T cell subsets (CD4+ and CD8+ cells), and macrophages in 2 and 8 month old Tff1-KO mice relative to Tff1-WT mice of matching age (Figure 3C-F, p<0.05). This increase in leukocytes, T lymphocytes subpopulations, and macrophages was significantly higher in older Tff1-KO (8 months) than younger Tff1-KO (2 months) mice (Figure 3B-F, p<0.01). The data also indicated that there was no significant change in immune cells infiltration in gastric tissues from Tff1-WT mice of all ages (Figure 3B-F).
Flow cytometry analysis for T lymphocytes and monocytes/macrophages in gastric tissue of Tff1-WT and Tff1-KO mice. (A) Representative flow cytometry dot plots showing gating strategy for gastric-infiltrating total leukocytes CD45+cells, total T lymphocytes CD45+CD3+cells, T cell subsets CD4+, CD8+, and monocytes/macrophages CD45+F4/80+cells. (B-F) Graphs showing flow cytometry data analysis indicating a significant increase of T lymphocytes and macrophages in Tff1-KO mice as compared to Tff1-WT (p<0.05). Notably, this increase was significantly greater in 8 months of age Tff1-KO mice than in 2-month-old mice (p<0.01). Data are expressed as mean ± SEM (n=3 to 5 mice per group).
Loss of Tff1 expression increases levels of T lymphocytes and monocytes/macrophages in mouse blood
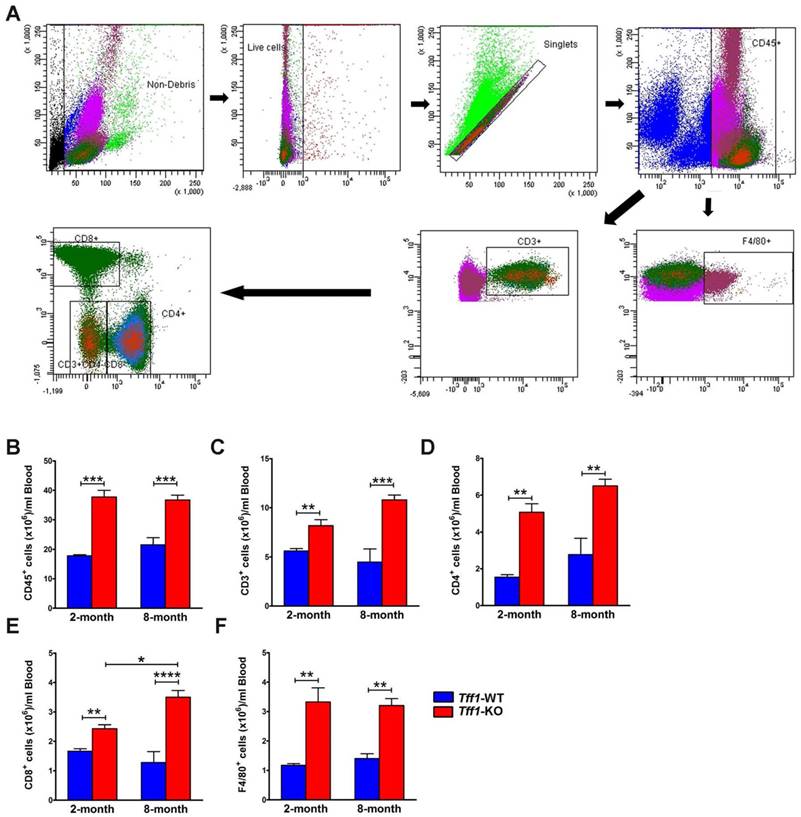
We investigated whether the levels of infiltrating immune cells in mouse gastric tissues were similar to those circulating in the blood stream. We performed flow cytometry to quantify total leukocytes (CD45+ cells), total T lymphocytes (CD45+CD3+ cells), T cell subsets (CD4+ and CD8+ cells), and monocytes/macrophages (CD45+F4/80+ cells) in blood samples from Tff1-KO and Tff1-WT mice (Figure 4A). We used 100 μl of blood collected from 2 and 8 month old mice immediately following their sacrifice. The data showed significant steady increases in all blood circulating immune cells in Tff1-KO in comparison with Tff1-WT mice of all ages (Figure 4B-F). These increases in circulating immune cells were generally similar in 2 and 8 month old Tff1-KO as compared to Tff1-WT mice (Figure 4B-F).
Tff1 deficiency increases levels of T lymphocytes and monocytes/macrophages in mouse blood. (A) Representative flow cytometry dot plots showing gating strategy for blood total leukocytes CD45+ cells, total T lymphocytes CD45+CD3+cells, T cell subsets CD4+, CD8+, and monocytes/macrophages CD45+F4/80+cells. (B-F) Graphs are showing flow cytometry data indicating a significant increase in T lymphocytes and macrophages levels in blood samples from Tff1-KO mice in comparison with Tff1-WT (p<0.01). Data are expressed as mean ± SEM (n=3 to 5 mice per group).
Loss of Tff1 expression increases levels of RORγt+ T lymphocytes and Il-17 mRNA in mouse gastric mucosa
Several reports indicated that activation of the oncogenic pathways, including NFκB and β-catenin, induces T lymphocytes differentiation by increasing the expression of transcription factor retinoic acid related orphan receptor-γt (RORγt) [36, 37]. Indeed, RORγt has been suggested to be a potential therapeutic target in chronic autoimmune and inflammatory diseases [38-40]. We evaluated the levels of RORγt+ cells in T lymphocytes, the progenitor cells that give rise to T cell subsets (CD3+CD4+ and CD3+CD8+ cells) in gastric tissues and blood from Tff1-KO and Tff1-WT mice. The flow cytometry data showed a significant increase in the ratio of RORγt+ T lymphocytes in gastric tissues from Tff1-KO, independent of age, as compared to Tff1-WT mice (Figure 5A, p<0.01). Conversely, in blood samples, there was no significant difference in RORγt+ T lymphocytes levels between Tff1-KO and Tff1-WT mice of matching age (Figure 5B). These data suggest that the increased levels of RORγt+ T lymphocytes in Tff1-KO relative to Tff1-WT mice are limited to gastric mucosa.
Loss of Tff1 expression increases CD3+RORγt+ T lymphocytes and Il-17 mRNA levels in gastric tissues. Graphs showing flow cytometry results of RORγt+CD3+T cells in gastric tissues (A) and Blood samples (B) from 2 and 8 month old Tff1-WT and Tff1-KO mice (n=3 to 5 mice per group). The data showed that the RORγt+CD3+T lymphocytes population was significantly higher in gastric tissues (p<0.01), but not in blood, in Tff1-KO than Tff1-WT mice of matching age. (C) QRT-PCR data indicating a significant increase of Il-17 mRNA expression (p<0.01) in mouse gastric tissues from 2 and 8 month old Tff1-KO as compared to Tff1-WT mice of matching age (n=10 mice per group).
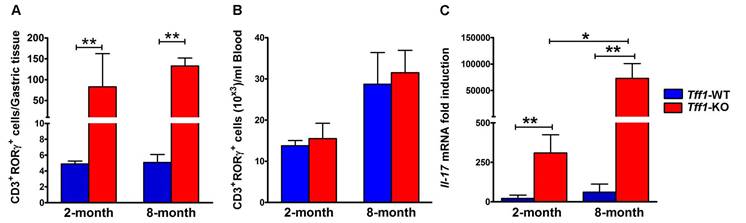
The RORγt nuclear receptor expressed by T lymphocytes drives the expression of pro-inflammatory cytokines such as IL-17 [18, Wang, 2012 #19]. IL-17 is an important cytokine secreted by T cells and other lymphoid cells have either a pro-tumor or an antitumor role in different cancer models [41-43]. Therefore, our next step was to evaluate the mRNA levels of Il-17 in gastric tissues from 2 and 8 month old Tff1-KO and Tff1-WT mice using real-time PCR analysis. The results revealed a significant increase in Il-17 mRNA levels in Tff1-KO, as compared to Tff1-WT of matching age (Figure 5C, p<0.01). Notably, Il-17 mRNA levels were significantly higher in 8-month-old than 2-month-old Tff1-KO mice (Figure 5C, p<0.05). These data indicate that the increase of inflammation in gastric tissues is associated with high levels of infiltrating RORγt+ T lymphocytes and Il-17 expression in Tff1-KO mice.
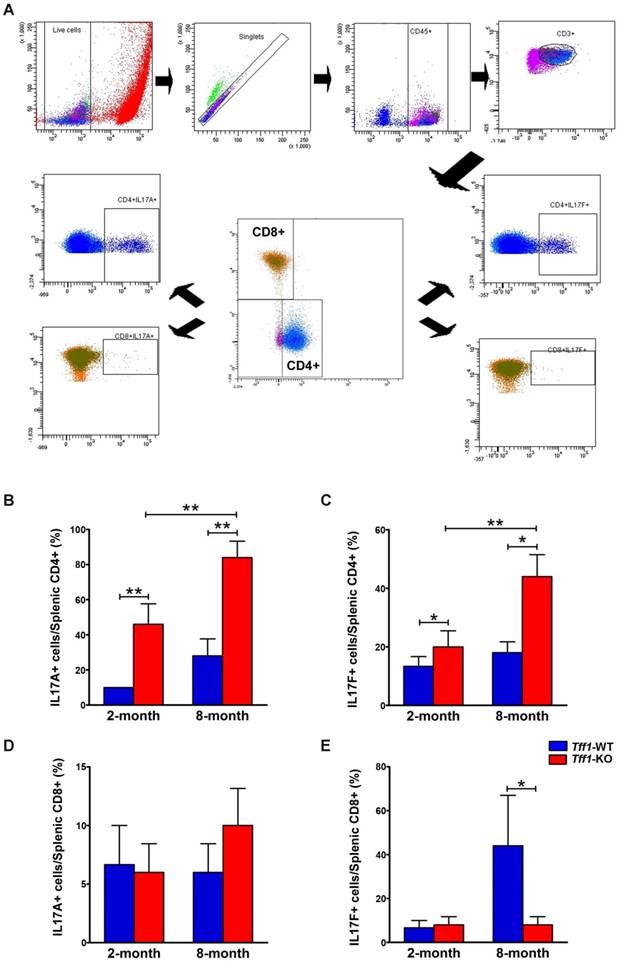
Loss of Tff1 promotes inflammation and suppresses cytotoxic response through regulation of Th17 and Tc17 cells
Th17 and Tc17 are different subsets of T lymphocytes derived from CD4+ and CD8+, respectively, and produce cytokines such as IL-17A and IL-17F [44]. To identify the population of IL-17-producing cells in our mouse model, spleen T lymphocytes were isolated from Tff1-WT and Tff1-KO mice at age 2 and 8 months and stimulated with PMA and ionomycin for 4 hours. Following intracellular staining, cells were analyzed by flow cytometry to determine the levels of Th17 and Tc17 subsets based on their expression of IL-17A and IL-17F. Our data indicated that levels of splenic Th17 cells expressing IL-17A and IL-17F were significantly higher in Tff1-KO than Tff1-WT mice of matching age (Figure 6B-C, p<0.05). In addition, the results showed that the increased levels of splenic Th17 cells were significantly higher in 8-month-old than 2-month-old Tff1-KO mice (Figure 6B-C, p<0.01). However, in the case of splenic cytotoxic Tc17 cells, derived from CD8+ cells, there was no significant change in the levels of IL-17A-positive cells in Tff1-KO as compared to Tff1-WT mice (Figure 6D), and a significant decrease of IL-17F-positive cells in 8-month-old Tff1-KO as compared to Tff1-WT mice of matching age (Figure 6E, p<0.05). Of note, the stimulation of the splenocytes significantly increased levels of IL-17F+ Tc17 cells in 8-month-old Tff1-WT, unlike Tff1-KO mice of matching age (Figure 6E, p<0.05). The data suggest that the increase of IL-17A+ and IL-17F+ in splenic Th17 cells derived from CD4+ cells, reflect the onset of an immune response in gastric tissues of the Tff1-KO mice, which promotes chronic inflammation and tumorigenesis. Interestingly, the decreased levels of IL-17F+ in splenic Tc17 cells in Tff1-KO in comparison to high levels of this cell population in Tff1-WT mice may suggest loss of the cytotoxic response in the Tff1-KO mice.
Loss of Tff1 increases the levels of IL-17A and IL-17F CD4+ but not in CD8+ cells splenocytes. (A) Representative gating strategy for the flow cytometry analysis of splenocytes isolated from 2 and 8 month old Tff1-KO and Tff1-WT mice (n=3 to 5 mice per group). Data analysis showing CD4+IL-17A+ lymphocytes (B), CD4+IL-17F+ lymphocytes (C), CD8+IL-17A+ lymphocytes (D), and CD8+IL-17F+ lymphocytes (E) in Tff1-KO and Tff1-WT mice of matching age. The data indicated a significant increase of CD4+IL-17A+ and CD4+IL-17F+ lymphocytes in Tff1-KO relative to Tff1-WT mice of matching age (p<0.01). There was no significant difference in CD8+IL-17A+ lymphocytes, but a significant decrease in CD8+IL-17F+ lymphocytes in Tff1-KO, as compared with Tff1-WT mice.
Discussion
Infiltration of immune cells in the tissue microenvironment and the onset of chronic inflammation play critical roles in initiation and promotion of tumor growth [5, 45]. Understanding the immune networks that exist within the tumor microenvironment is essential for not only understanding the tumorigenesis cascade but also for designing and developing a potential therapeutic strategy. In this study, we characterize the distribution of infiltrating immune cells after Tff1 loss in gastric mouse tissue at early and late stage of tumor formation.
We previously demonstrated that TFF1, a secreted protein in the stomach, has an anti-inflammatory effect where the Tff1-KO mouse model develops gastric tumors through activation of NFκB and chronic inflammatory pathways [23]. In this report, we confirmed this finding and provided evidence that loss of Tff1 provides a perfect setting for infiltration of immune cells in the tissue microenvironment, which subsequently promotes chronic inflammation and tumorigenesis. Our data indicated that infiltration of immune cells in the gastric mucosa of the Tff1-KO mouse were age-dependent progressive changes that were present at a young age and early stages of gastric tumorigenesis.
Several studies have shown that the tumor microenvironment has increased infiltration of inflammatory cells, particularly lymphocytes and macrophages [46, 47]. Our immunohistochemistry analysis of gastric tissues demonstrated increased infiltration of CD3 positive lymphocytes and F4/80 positive macrophages in histological lesions of the Tff1-KO mouse. These findings confirm the inflammatory phenotype after loss of Tff1, and support our flow cytometry data showing an increase of immune cell populations such as leukocytes (CD45+), T lymphocytes (CD45+ CD3+), T cell subsets (CD4+, CD8+) and Monocytes/Macrophages (CD45+ F4/80) in mouse gastric tissues. Indeed, our results demonstrated that this increase was also age dependent by showing more expression of these cells at the age of 8 months, a finding that confirms the mutual relationship between aging, inflammation and chronic diseases such as cancer [48, 49]. Although analysis of blood samples, the recruiter source of the immune cell to the tumor microenvironment, showed an increase of immune cell markers in the Tff1-KO mouse model, this increase was not age-dependent. This finding suggests that the increased recruitment of immune cells in old age Tff1-KO mice could be related to the progressive nature of histological lesions with the occurrence of high grade dysplasia and invasive adenocarcinoma at old age in these mice.
In gastric cancer, earlier reports have demonstrated an increase of the transcription factor retinoic acid related orphan receptor-γt (RORγt) especially in metastatic patients who are characterized by poor clinical outcome [50, 51]. Moreover, the activation of oncogenic pathways, such as NFκB and β-catenin, has been shown to induce regulation of T cell differentiation by regulating the expression of RORγt [36, 37]. In our previous reports, we found that loss of Tff1 promotes activation of NFκB and β-catenin pathways in mouse and human gastric neoplasms [23, 25]. Herein, our data confirmed that loss of Tff1 induces an increase of T cell RORγt positive cells in mouse gastric tissues, not in blood samples, suggesting that the changes in T cell expression are tissue specific.
CD4+T cells, which are key regulators of the immune system, differentiate into T helper (Th17) cell lineages that can promote chronic inflammation and tumor growth [52, 53]. CD8+ cells are the next subset of T cells that can differentiate into Tc17 and exhibit strong cytotoxic activity in various conditions, such as infection and cancer [54, 55]. Th17 and Tc17 activating T regulatory cells subsets, both express RORγt marks and have the potential to express cytokines such as IL-17A and IL-17F [18, 56].
Our data are in agreement with the aforementioned reports by demonstrating the pro-inflammatory effect of Th17. Indeed, we have demonstrated an increase of IL-17A and IL-17F in CD4+ (Th17) indicating the inflammatory phenotype observed in Tff1-KO mice that was also found to be age-dependent. These data were confirmed by examining mRNA expression levels of IL-17 in mouse gastric tissues, which were significantly higher in Tff1-KO as compared to Tff1-WT. However, the flow cytometry analysis of CD8+ cells showed no changes in IL-17A in Tff1-KO gastric tissues as compared to Tff1-WT. Nonetheless, we noticed a decrease of IL-17F at the age of 8 months in gastric tissue of Tff1-KO compared to Tff1-WT of matching age. These results are in agreement with previous reports showing a protective effect of IL-17F in colonic epithelial cells against tumorigenesis [57]. Therefore, this decrease in IL-17F could be due to loss of cytotoxic effect of the CD8+ population and the development of gastric tumors in our mouse model.
In summary, gastric cancer cells can bypass the immune surveillance by suppressing the immune system and inducing immunological tolerance. The stroma and tumor microenvironment of gastric tissues in the Tff1-KO mouse model revealed strong evidence supporting the presence of T cell-based inflammation and loss of the antitumor cytotoxic T cells. Future studies are needed to investigate mechanisms by which secreted gastric Tff1 protein regulates interactions and mutual regulation of these complex immune networks within the tumor microenvironment in gastric cancer.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Weber C, Telerman SB, Reimer AS, Sequeira I, Liakath-Ali K, Arwert EN. et al. Macrophage infiltration and alternative activation during wound healing promote MEK1-induced skin carcinogenesis. Cancer Res. 2016
2. Chew V, Toh HC, Abastado JP. Immune microenvironment in tumor progression: characteristics and challenges for therapy. J Oncol. 2012;2012:608406
3. Shiao SL, Ganesan AP, Rugo HS, Coussens LM. Immune microenvironments in solid tumors: new targets for therapy. Genes Dev. 2011;25:2559-72
4. Dagenais M, Dupaul-Chicoine J, Champagne C, Skeldon A, Morizot A, Saleh M. A critical role for cellular inhibitor of protein 2 (cIAP2) in colitis-associated colorectal cancer and intestinal homeostasis mediated by the inflammasome and survival pathways. Mucosal Immunol. 2016;9:146-58
5. Landskron G, De la Fuente M, Thuwajit P, Thuwajit C, Hermoso MA. Chronic inflammation and cytokines in the tumor microenvironment. J Immunol Res. 2014;2014:149185
6. Karin M, Greten FR. NF-kappaB: linking inflammation and immunity to cancer development and progression. Nat Rev Immunol. 2005;5:749-59
7. Balkwill F, Charles KA, Mantovani A. Smoldering and polarized inflammation in the initiation and promotion of malignant disease. Cancer Cell. 2005;7:211-7
8. Karin M, Lawrence T, Nizet V. Innate immunity gone awry: linking microbial infections to chronic inflammation and cancer. Cell. 2006;124:823-35
9. Gajewski TF, Schreiber H, Fu YX. Innate and adaptive immune cells in the tumor microenvironment. Nat Immunol. 2013;14:1014-22
10. de Visser KE, Eichten A, Coussens LM. Paradoxical roles of the immune system during cancer development. Nat Rev Cancer. 2006;6:24-37
11. DeNardo DG, Johansson M, Coussens LM. Immune cells as mediators of solid tumor metastasis. Cancer Metastasis Rev. 2008;27:11-8
12. Moseley TA, Haudenschild DR, Rose L, Reddi AH. Interleukin-17 family and IL-17 receptors. Cytokine Growth Factor Rev. 2003;14:155-74
13. Lockhart E, Green AM, Flynn JL. IL-17 production is dominated by gammadelta T cells rather than CD4 T cells during Mycobacterium tuberculosis infection. J Immunol. 2006;177:4662-9
14. Liu SJ, Tsai JP, Shen CR, Sher YP, Hsieh CL, Yeh YC. et al. Induction of a distinct CD8 Tnc17 subset by transforming growth factor-beta and interleukin-6. J Leukoc Biol. 2007;82:354-60
15. Zhong F, Cui D, Tao H, Du H, Xing C. IL-17A-producing T cells and associated cytokines are involved in the progression of gastric cancer. Oncol Rep. 2015;34:2365-74
16. Liu J, Xu Q, Yuan Q, Wang Z, Xing C, Yuan Y. Association of IL-17A and IL-17F polymorphisms with gastric cancer risk in Asians: a meta-analysis. Hum Immunol. 2015;76:6-12
17. Garcia-Hernandez Mde L, Hamada H, Reome JB, Misra SK, Tighe MP, Dutton RW. Adoptive transfer of tumor-specific Tc17 effector T cells controls the growth of B16 melanoma in mice. J Immunol. 2010;184:4215-27
18. Ivanov II, McKenzie BS Zhou L, Tadokoro CE Lepelley A, Lafaille JJ et al. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell. 2006;126:1121-33
19. Wang X, Zhang Y, Yang XO, Nurieva RI, Chang SH, Ojeda SS. et al. Transcription of Il17 and Il17f is controlled by conserved noncoding sequence 2. Immunity. 2012;36:23-31
20. Thim L, May FE. Structure of mammalian trefoil factors and functional insights. Cell Mol Life Sci. 2005;62:2956-73
21. Lefebvre O, Chenard MP, Masson R, Linares J, Dierich A, LeMeur M. et al. Gastric mucosa abnormalities and tumorigenesis in mice lacking the pS2 trefoil protein. Science. 1996;274:259-62
22. Buache E, Etique N, Alpy F, Stoll I, Muckensturm M, Reina-San-Martin B. et al. Deficiency in trefoil factor 1 (TFF1) increases tumorigenicity of human breast cancer cells and mammary tumor development in TFF1-knockout mice. Oncogene. 2011;30:3261-73
23. Soutto M, Belkhiri A, Piazuelo MB, Schneider BG, Peng D, Jiang A. et al. Loss of TFF1 is associated with activation of NF-kappaB-mediated inflammation and gastric neoplasia in mice and humans. J Clin Invest. 2011;121:1753-67
24. Soutto M, Chen Z, Saleh MA, Katsha A, Zhu S, Zaika A. et al. TFF1 activates p53 through down-regulation of miR-504 in gastric cancer. Oncotarget. 2014;5:5663-73
25. Soutto M, Peng D, Katsha A, Chen Z, Piazuelo MB, Washington MK. et al. Activation of beta-catenin signalling by TFF1 loss promotes cell proliferation and gastric tumorigenesis. Gut. 2015;64:1028-39
26. Soutto M, Romero-Gallo J, Krishna U, Piazuelo MB, Washington MK, Belkhiri A. et al. Loss of TFF1 promotes Helicobacter pylori-induced beta-catenin activation and gastric tumorigenesis. Oncotarget. 2015;6:17911-22
27. Beckler AD, Roche JK, Harper JC, Petroni G, Frierson HF Jr, Moskaluk CA. et al. Decreased abundance of trefoil factor 1 transcript in the majority of gastric carcinomas. Cancer. 2003;98:2184-91
28. Park WS, Oh RR, Park JY, Lee JH, Shin MS, Kim HS. et al. Somatic mutations of the trefoil factor family 1 gene in gastric cancer. Gastroenterology. 2000;119:691-8
29. Carvalho R, Kayademir T, Soares P, Canedo P, Sousa S, Oliveira C. et al. Loss of heterozygosity and promoter methylation, but not mutation, may underlie loss of TFF1 in gastric carcinoma. Lab Invest. 2002;82:1319-26
30. Tomita H, Takaishi S, Menheniott TR, Yang X, Shibata W, Jin G. et al. Inhibition of gastric carcinogenesis by the hormone gastrin is mediated by suppression of TFF1 epigenetic silencing. Gastroenterology. 2011;140:879-91
31. Katsha A, Soutto M, Sehdev V, Peng D, Washington MK, Piazuelo MB. et al. Aurora kinase A promotes inflammation and tumorigenesis in mice and human gastric neoplasia. Gastroenterology. 2013;145:1312-22 e1-8
32. El-Rifai W, Moskaluk CA, Abdrabbo MK, Harper J, Yoshida C, Riggins GJ. et al. Gastric cancers overexpress S100A calcium-binding proteins. Cancer Res. 2002;62:6823-6
33. Coussens LM, Werb Z. Inflammation and cancer. Nature. 2002;420:860-7
34. Smyth MJ, Cretney E, Kershaw MH, Hayakawa Y. Cytokines in cancer immunity and immunotherapy. Immunol Rev. 2004;202:275-93
35. Khong HT, Restifo NP. Natural selection of tumor variants in the generation of "tumor escape" phenotypes. Nat Immunol. 2002;3:999-1005
36. Keerthivasan S, Aghajani K, Dose M, Molinero L, Khan MW, Venkateswaran V. et al. beta-Catenin promotes colitis and colon cancer through imprinting of proinflammatory properties in T cells. Sci Transl Med. 2014;6:225ra28
37. Oh H, Ghosh S. NF-kappaB: roles and regulation in different CD4(+) T-cell subsets. Immunol Rev. 2013;252:41-51
38. Withers DR, Hepworth MR, Wang X, Mackley EC, Halford EE, Dutton EE. et al. Transient inhibition of ROR-gammat therapeutically limits intestinal inflammation by reducing TH17 cells and preserving group 3 innate lymphoid cells. Nat Med. 2016;22:319-23
39. Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 Cells. Annu Rev Immunol. 2009;27:485-517
40. Gaffen SL, Jain R, Garg AV, Cua DJ. The IL-23-IL-17 immune axis: from mechanisms to therapeutic testing. Nat Rev Immunol. 2014;14:585-600
41. Res PC, Piskin G, de Boer OJ, van der Loos CM, Teeling P, Bos JD. et al. Overrepresentation of IL-17A and IL-22 producing CD8 T cells in lesional skin suggests their involvement in the pathogenesis of psoriasis. PLoS One. 2010;5:e14108
42. Murugaiyan G, Saha B. Protumor vs antitumor functions of IL-17. J Immunol. 2009;183:4169-75
43. Cua DJ, Tato CM. Innate IL-17-producing cells: the sentinels of the immune system. Nat Rev Immunol. 2010;10:479-89
44. Iwakura Y, Ishigame H, Saijo S, Nakae S. Functional specialization of interleukin-17 family members. Immunity. 2011;34:149-62
45. Zamarron BF, Chen W. Dual roles of immune cells and their factors in cancer development and progression. Int J Biol Sci. 2011;7:651-8
46. Mahmoud SM, Paish EC, Powe DG, Macmillan RD, Grainge MJ, Lee AH. et al. Tumor-infiltrating CD8+ lymphocytes predict clinical outcome in breast cancer. J Clin Oncol. 2011;29:1949-55
47. D'Angelo SP, Shoushtari AN, Agaram NP, Kuk D, Qin LX, Carvajal RD. et al. Prevalence of tumor-infiltrating lymphocytes and PD-L1 expression in the soft tissue sarcoma microenvironment. Hum Pathol. 2015;46:357-65
48. Goubran HA, Kotb RR, Stakiw J, Emara ME, Burnouf T. Regulation of tumor growth and metastasis: the role of tumor microenvironment. Cancer Growth Metastasis. 2014;7:9-18
49. Ahmad A, Banerjee S, Wang Z, Kong D, Majumdar AP, Sarkar FH. Aging and inflammation: etiological culprits of cancer. Curr Aging Sci. 2009;2:174-86
50. Su Z, Sun Y, Zhu H, Liu Y, Lin X, Shen H. et al. Th17 cell expansion in gastric cancer may contribute to cancer development and metastasis. Immunol Res. 2014;58:118-24
51. Yoshida N, Kinugasa T, Miyoshi H, Sato K, Yuge K, Ohchi T. et al. A High RORgammaT/CD3 Ratio is a Strong Prognostic Factor for Postoperative Survival in Advanced Colorectal Cancer: Analysis of Helper T Cell Lymphocytes (Th1, Th2, Th17 and Regulatory T Cells). Ann Surg Oncol. 2016;23:919-27
52. Bailey SR, Nelson MH, Himes RA, Li Z, Mehrotra S, Paulos CM. Th17 cells in cancer: the ultimate identity crisis. Front Immunol. 2014;5:276
53. Lenoir M, Del Carmen S, Cortes-Perez NG, Lozano-Ojalvo D, Munoz-Provencio D, Chain F. et al. Lactobacillus casei BL23 regulates T and Th17 T-cell populations and reduces DMH-associated colorectal cancer. J Gastroenterol. 2016
54. Liang Y, Pan HF, Ye DQ. Tc17 Cells in Immunity and Systemic Autoimmunity. Int Rev Immunol. 2015;34:318-31
55. Won TJ, Lee YJ, Hyung KE, Yang E, Sohn UD, Min HY. et al. SUMO2 overexpression enhances the generation and function of interleukin-17-producing CD8(+) T cells in mice. Cell Signal. 2015;27:1246-52
56. Blatner NR, Mulcahy MF, Dennis KL, Scholtens D, Bentrem DJ, Phillips JD. et al. Expression of RORgammat marks a pathogenic regulatory T cell subset in human colon cancer. Sci Transl Med. 2012;4:164ra59
57. Tong Z, Yang XO, Yan H, Liu W, Niu X, Shi Y. et al. A protective role by interleukin-17F in colon tumorigenesis. PLoS One. 2012;7:e34959
Author contact
![]() Corresponding author: Wael El-Rifai, M.D., Ph.D., Vanderbilt University Medical Center, 760 Preston Research Bldg., 2220 Pierce Avenue, Nashville, TN 37232 E-mail: wael.el-rifaiedu, Phone: 615-322-7934, Fax: 615-322-7852
Corresponding author: Wael El-Rifai, M.D., Ph.D., Vanderbilt University Medical Center, 760 Preston Research Bldg., 2220 Pierce Avenue, Nashville, TN 37232 E-mail: wael.el-rifaiedu, Phone: 615-322-7934, Fax: 615-322-7852