Impact Factor ISSN: 1837-9664

 Global reach, higher impact
Global reach, higher impactJ Cancer 2026; 17(7):1384-1392. doi:10.7150/jca.135857 This issue Cite
Review
tsRNA's Biological Function and its Potential Application in Disease Diagnosis and Prognosis
Congcong Tang1,2*, Qi Tang1*, Siyuan Tang2, Faqing Tang1 ![]()
1. Hunan Key Laboratory of Oncotarget Gene, Hunan Cancer Hospital, and The Affiliated Cancer Hospital of Xiangya School of Medicine, Central South University, Changsha 410013, China.
2. Xiangya School of Nursing, Central South University, Changsha, China.
* These authors have equal contribution.
Received 2026-4-8; Accepted 2026-6-30; Published 2026-7-14
Abstract

Under physiological conditions, tsRNAs regulate mRNA transcription, reverse transcription, and protein translation to mediate cell apoptosis, cell cycle, and epigenetic regulation. Pathogenic tsRNAs are disease-specific or disease-related tsRNAs that are highly express in various diseases and may serve as diagnostic or potential diagnostic biomarkers. Specifically expressed tsRNAs in tumors were screened and obtained from the tumors. Although the mechanisms by which these oncogenic tsRNAs contribute to tumor development remain unclear, they have been shown to be applicable to tumor diagnosis and therapeutic prognosis. This article briefly summarizes pathogenic tsRNAs involved in various diseases and their biological functions. Oncogenic tsRNAs in tumors and their clinical applications have been elaborated upon. The molecular mechanisms of pathological tsRNAs in both general diseases and tumors need to be further investigated in future.
Keywords: tsRNA, regulatory mechanism, biomarker, non-tumor disease, tumor
Introduction
tRNA is a fundamental component of the translation mechanism that delivers amino acids to ribosomes and translates genetic information into corresponding peptide chains. It is the most abundant of all non-coding RNA small molecules, accounting for 4-10% of all cellular RNA [1]. Aminoacyl-tRNA is activated, accurately transferring amino acids to the peptide chain. tsRNAs are small RNAs derived from tRNAs and also known as tRNA-derived fragments (tRF), tRNA-derived stress-induced RNAs (tiRNAs), and semi-tRNA. tsRNAs are a recently discovered class of small non coding RNAs, produced by the cleavage of mature tRNA or tRNA precursors by enzymes, such as angiogenin (ANG), Dicer, RNase Z, and RNase P [2]. tRNA genes are transcribed into precursor tRNAs (pre-tRNAs) through RNA polymerase III (RNA Pol III), each of which has a 5'leading sequence and a 3' tail region [3]. Usually, RNase P removes the leading segment to the 5'end [4], and RNase Z removes the trailing segment at the 3' end [5]; nucleotide transferase adds the “CCA” sequence to the 3 'end [6]; subsequently the sequence is folded into the secondary clover structure of mature tRNA by post transcriptional modification. The structure of tRNA clover consists of a dihydrouracil ring (D ring), dihydrouracil arm, anticodon ring, anticodon arm, variable ring, pseudouracil ring (T ψ C ring or T ring), pseudouracil arm, and amino acid arm [3]. tsRNAs exert biological functions through various mechanisms, such as interacting with proteins or mRNA, inhibiting translation, and regulating gene expression, cell cycle, chromatin, and epigenetic modifications [7,8]. Recently, some disease-specific and disease-related tsRNAs have been discovered [9-14], and are known as pathogenic tsRNA. They have been verified to play critical biological functions in the occurrence and development of diseases [15,16], and some tumor-specific and tumor-related tsRNAs have been screened and applied in tumor diagnosis and therapy [17-22]. This article systematically expounds tsRNA formation mechanisms and biofuncation. We also briefly summarized the tsRNAs applications of tsRNA in disease diagnosis. In particular, the diagnostic application and therapeutic prognosis of patients with tumors are importantly elaborated.
1. Formation mechanisms of tsRNA and biological functions
tsRNAs include i-tRF, tRF-1, tRF-2, tRF-3, tRF-5, tiRNA, sitRNA-5, and sitRNA-3 [23-25]. tRFs and tiRNAs are a class of small non-coding RNAs produced by site-specific cleavage of mature or precursor tRNAs, but their nomenclature remains inconsistent. Collectively, termed tRNA-derived small RNAs or tRNA-derived RNAs, these molecules are known by multiple interchangeable names, including tRFs, tiRNAs, tRNA halves, SHOT-RNAs, and tsncRNAs. The most widely adopted classification divides them into two main groups, longer tiRNAs (29-50 nt) and shorter tRFs (14-30 nt). The former is also called stress-induced tRNA halves, which are generated by angiogenin-mediated cleavage within the anticodon loop into 5′-tiRNA and 3′-tiRNA. The latter is further subdivided according to their origins, tRF-5 from the 5′ end (cleaved at the D-loop), tRF-3 from the 3′ end (cleaved at the T-loop, containing CCA motif), tRF-1 from the 3′ trailer of pre-tRNAs (with a poly-U tail), and internal i-tRFs spanning central regions. In addition, the databases of tRFdb use numerical IDs to name tRF, such as 5001 for tRF-5 and 3001 for tRF-3, while other systems incorporate tRNA isotype, amino acid, anticodon, or genomic position. Some research groups adopt distinct naming conventions based on fragment length (5′ tRF, 3′ tRF, and i-tRF), processing origin (tRF-1, tRF-2, tRF-5a/b/c), or the specific tRNA isodecoder from which they originate, such as 5′ tRFs derived from the 5′ end of mature tRNAs, 3′ tRFs from the 3′ end, and internal tRFs (i-tRFs) from internal regions are commonly described [26,27].
i-tRF, tRF-2, tRF-3, and tRF-5 are derived from mature tRNAs through cleavage by ANG [28] and Dicer [29], or from other tRNAs that RNase cleavages RNA at different sites of RNA. tRF-1s are derived from pre-tRNAs cleaved by RNase Z decomposing. tiRNAs are also classified into two types, 5'tiRNA and 3'tiRNA. They are derived from mature tRNAs that are cleaved from the anticodon loop. A fragment longer than the RNA is called stress-induced sitRNA-5 or sitRNA-3 [30]. tRF-5 is usually present in the nucleus [31], whereas tRF-1 and tRF-3 are mainly present in the cytoplasm [32,33]. Methylation [34] is the most common modification of tsRNA, and some tRNAs that contain 5'-terminal oligoguanine can regulate the expression of 5'- tRFs through pseudouridylation [35]. Nucleoside modification [36] also plays important roles in tsRNA biology. The main biological functions of tsRNA are described as follows.
1.1 tsRNA regulates mRNA transcription
Recent studies have shown that tsRNAs recognize target mRNAs and bind to them during transcription, when the untranslated region is replaced and the transcriptional stability is decreased, thereby inhibiting mRNA expression [37]. Zhang et al. discovered a 5'-tRF, which is generated from piRNA (td piR (Glu)) that is derived from 5'-tRNAGlu. 5'-tRF was proved to bind to PIWI-like protein 4 and recruit SET domain bifurcated histone lysine methyltransferase 1, suppressor of variegation 39 homolog 1, and heterochromatin protein 1 β to the cluster of differentiation 1A (CD1A) promoter region, thereby which regulates H3K9 methylation and inhibits human monocyte CD1A transcription [38]. Chen et al. [39] suggested that sperm tsRNAs interact with promoter regions, but not coding regions, promoting downstream gene expression, and that binding alters the inheritance of metabolic traits and embryonic development. Collectively, stRNAs have respective and intricate molecular mechanisms to participate into cellular biological behavior, stRNAs from different origins regulate gene transcriptions via different mechanism. A group of i-tRFs from tRNAGlu, tRNAAsp, tRNAGly, and tRNATyr bind to the 3 'end of multiple oncogenic transcripts through competing with Y-box binding protein 1 (YB1), which results in a decrease in expression of target oncogenes, thus inhibiting breast cancer metastasis [40]. In addition, tsGlnCTG interacts with insulin-like growth factor 2 mRNA-binding protein 1 to inhibit transcript stability and promote stem cell differentiation [41]. 3'tsRNAHisGTG and 3'tsRNALeuCAG exert crucial roles in gene expressions, 17nt 3'tsRNAHisGTG and 18nt 3'tsRNALeuCAG induce the cleavage of artificial target mRNA mediated by endogenous argonaute (AGO)2 [42]. AGO family members are important tsRNA targets. Kumar P et al. found that tsRNAs associated with AGOs regulate the gene silencing [31], and further discovered that type I tsRNAs interact more easily with AGO1, AGO3, and AGO4 than with AGO2, whereas type II tsRNAs have almost no interaction with AGO protein. Maute et al. [43] evaluated the interaction of CU1276tsRNA (3'tsRNAGlyGCC) with AGO1-4 proteins through immunoprecipitation experiments and found that these interactions repress mRNA transcripts in a sequence-specific manner. The relationship between tsRNAs and AGO protein was verified in fruit flies, and the results showed that the interaction of tsRNAs with AGO1 or AGO2 changes with age [44]. The downstream molecules of tsRNAs have some specificity, various AGO-interacting tsRNAs interact with different AGO family members, tsRNA function is determined to some extent by its downstream AGO members.
1.2 tsRNA regulates protein translation
tRFs and tiRNA may inhibit protein translation by disrupting ribosome formation and prolonging protein activity [45]. Under specific situation, halophilic archaeon Haloferax volcanii from tsRNAs produced by tRNAVal5 fragments inhibited peptidyl transferase activity and weakened translation by binding to small nucleosome subunits [46]. Arabidopsis can bind to multiple ribosomes to generate tRFs that inhibit plant translation [47]. Moreover, 5'-iRNAAla and 5'-iRNACys replace the cap end of the complex eukaryotic initiation factor gamma 4F (eIF4G/A/F) by binding to the translation inhibitory factor YB-1 to induce the assembly of stress granules, thus inhibiting translation initiation [48]. Subsequently, G-quadruplex 4 tiRNA was shown to bind directly to the HEAT1 domain of eIF4G, resulting in impaired 40S ribosome binding and translation silencing [49]. Another report proved that deletion of cytosin-5 RNA methyltransferase NSun2 leads to an accumulation of 5'- tRFs and a decrease in protein translation rate [50]. Pseudouridylate synthase 7 mediated tRF-5 pseudouridylation can inhibit stem cell translation [35]. tsRNA5GluCTC mediated by respiratory syncytial virus (RSV) promotes RSV replication, which reduces RSV defense genes expression and tsRNA5GluCTC interaction with RSV proteins [51]. The silencing complex formed through tRF-3 binding to Ago3 and Ago4 directly binds to the mRNA of the target gene, ultimately inhibiting its translation of target gene. It was found that 5'-tRFGln19 interacts with triple synthase complex, promoting ribosome binding to poly (A) tail and translation elongation [52].
1.3 tsRNA mediates reverse transcription
Some tsRNAs regulate reverse transcription, such as the tRFs originating from the front tail region of tRNA, which regulates viral gene expression by isolating La/SSB from the cytoplasm [53]. tRF mediated by tRNAGlyGCC inhibits nearly 70 endogenous reverse transcription factor MERVL related genes transcription [56]. RSV infection induces tsRNA5GlyCTC, tsRNA5GlyGCC, tsRNAF5LysCTT, and tsRNA5CysGCA [51,57], tsRNA5GlyCTC mediates RSV replication by inhibiting apolipoprotein E receptor 2 gene transcription [58]. tsRNAs targeting the primer binding sites (PBS) of tRNA mediate the inhibition of gene transcription. Moreover, 18 nt 3'-tRF from dsRNA inhibited HIV-1 reverse transcription through targeting PBS, suppressing HIV-1 proliferation [54]. Conversely, tRF-3019 binds to PBS as a primer for the human T-cell leukemia virus type 1 reverse transcriptase, enhancing viral infection [55].
1.4 tsRNA is involved in cell apoptosis and cell cycle regulation
Thyroid papillary carcinoma (PTC) detection found that 33 nt 5'tiRNAGlyGCC is elevated in PTC, and it binds to the U2AF homology motif domain of RNA binding motif protein 17 (RBM17) and promotes RBM17 to translocate into the nucleus. Stable expressed-RBM17 induces alternative splicing of mitogen-activated protein kinase 4 mRNA exon 16, leading to proliferation and migration of PTC cells [59]. The immunohistochemistry detection showed that tsRNA-26576 is highly expressed in breast cancer tissue, and the cell experiments have shown that tsRNA-26576 inhibits cell apoptosis and induces the proliferation and migration of MDA-MB-231 cells [60]. 3'tsRNAValCAC2 directly binds to partner molecule eukaryotic translation elongation factor 1α1 (EEF1A1) and mediates its transport to the nucleus, thereby promoting the interaction with p53 E3 ubiquitin ligase MDM2. This indicates that 3'tsRNAValCAC2 inhibits gastric cancer cell apoptosis via the MDM2-p53 pathway, resulting in cancer progression [61]. In mouse models, knock-down of 3'tsRNALeuCAG induced significant apoptosis in HeLa cells, and decreased cancer xenografts derived from patients [62]. In wild-type mouse embryonic fibroblasts (MEF), high osmotic stress induces the mitochondrial release of cytochrome c (Cyt c) and subsequent formation of apoptotic bodies, finally leading to cell apoptosis. Mechanically, stress granules isolate Cyt C in ribonucleoprotein complexes by inducing 5'- and 3'- tiRNAs, thereby protecting MEF or primary neurons from osmotic stress, inducing apoptosis [63]. tRF-1001 is a type of tsRNA-1 derived from the 3'end of tRNASer precursor, it is essential for tumor cell growth [64]. Knock-down of tRF-1001 causes cells to remain in the G2 phase by inhibiting DNA biosynthesis, which interferes with cell proliferation [65]. Some tRFs and tiRNAs bind to cytochrome C, inhibiting the interaction of cytochrome C with Apaf-1 to block the activation of caspase-9 and restrain apoptotic body formation, thus suppressing apoptosis [63,66]. In addition, tRNA methyltransferase 10A mediated by 5'tRNAGln can induce pancreatic beta cell death [67].
1.5 tsRNAs in the epigenetic inheritance of acquired traits
Some tRFs from the sperm or epigenetic factors induce the expression of certain offspring genes, causing metabolic disorders. tsRNAs have been reported to act as epigenetic regulators that affect offspring metabolism. Male mice fed a high-fat diet (HFD) developed insulin resistance and impaired glucose tolerance, which occurred 7 weeks after birth and became severe at 15 weeks. This finding was verified to be caused by 30nt-34nt tRFs [68]. The high-fat diet of parental mice altered the expression profile of specific tsRNAs (e.g., 5'-tRFGlyGCC) in sperm, and the injection of these tsRNAs into normally fertilized eggs resulted in similar metabolic abnormalities (e.g., insulin resistance) in offspring. This is direct evidence that environmental factors (diet) can pass on acquired metabolic traits to offspring by altering tsRNA profiles in reproductive cells without altering DNA sequences. In addition, tRFGluTTC is considered a novel epigenetic regulator that promotes fat production [69]. One finding demonstrates that tRFGly functions as a key regulator of fat accumulation, targeting tRFGly may represent a therapeutic strategy for related metabolic disorders [70]. Moreover, this mechanism may be involved the regulation of mitochondrial tsRNA in fat deposition, suggesting that tsRNA may affect the intergenerational inheritance of energy metabolism by regulating mitochondrial function. Another study in the mice fed a low-protein diet confirmed that tsRNAs play critical roles in mammalian sperm maturation and fertilization. A low-fat diet was not associated with tsRNAs in immature sperm from mouse testes but had a significant effect on tsRNAs in mature sperm, particularly with a significant increase in tRNAGlyGCC levels [56]. As a RNA modification enzyme, DNMT2 plays a role in this process by changing the modification pattern of tsRNAs [71]. The latest research shows that the microRNAs carried by sperm (including tsRNA) not only affect fertilization rate, but also directly determine embryo quality and developmental potential [72]. In addition, tRFGluTTC is considered a novel epigenetic regulator that promotes fat production [73].
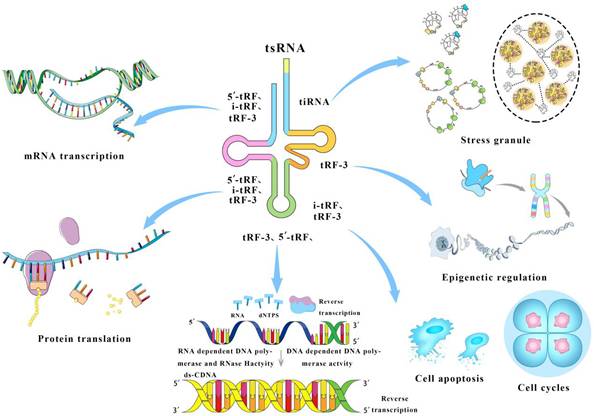
Biological functions of tsRNAs are summarized in Figure. Briefly, 5'-tRF, i-tRF, and tRF-3 respectively recognize their respective target molecules, inhibiting mRNA expression. 5'-tRF, i-tRF, and tRF-3 inhibit protein translation by disturbing ribosome formation and prolong protein activity. tRF-3 and 5'-tRFs regulate reverse transcription through interaction with reverse transcriptase. i-tRF and tRF-3 regulate cell cycle process via DNA biosynthesis, and participate in cell apoptosis through activating caspase-9 and restraining apoptotic body formation. tRF-3 participates in epigenetic inheritance via affecting the metabolism of offspring. tiRNAs including 5'- and 3'-tiRNAs, induce stress granules to isolate Cyt C in ribonucleoprotein complexes, thereby protecting MEF or primary neurons from osmotic stress.
Function and mechanism Schematic of tsRNAs participating in various biological activities. 5'-tRF, i-tRF, and tRF-3 mediate the inhibition of mRNA transcription. 5'-tRF, i-tRF, and tRF-3 inhibit protein translation. tRF-3 and 5'-tRFs regulate reverse transcription through interaction with reverse transcriptase. i-tRF and tRF-3 regulate cell cycle process and participate in cell apoptosis. tRF-3 participates in epigenetic inheritance. tiRNAs induce stress granules.
2. tsRNAs from patients with various tumors and clinical significance
2.1 tsRNAs are applied in tumor diagnosis
Serum tRFProAGG004 and tRFLeuCAG002 levels can be used as new biomarkers for pancreatic cancer diagnosis, even in the early stage [74]. In addition, tsRNAValTAC41 and tsRNAMetCAT37 were validated to have diagnostic value in patients with pancreatic ductal adenocarcinoma [75]. Compared with normal controls, tRF-5 derived from tRNAHis is downregulated in chronic lymphocytic leukemia [76]. tsRNA-46 and tsRNA-47 are strongly downregulated tsRNAs in chronic lymphocytic leukemia and lung cancer [77]. tsRNA-40 is upregulated in colon cancer, ts-66 and ts-86 are significantly dysregulated in breast cancer, and tsRNA-29 is overexpressed in ovarian cancer [77]. More interestingly, tRFGlnTTG006 can distinguish hepatocellular carcinoma from healthy individuals with high sensitivity (80.4%) and specificity (79.4%), even in stage I with a sensitivity of 79.0% and specificity of 74.8% [78]. In addition, the levels of tRNA-5 in plasma exosomes from patients with liver cancer were significantly higher than those in healthy controls [79]. Shen et al. suggested that tRF-33-P4R8YP9LON4VDP is a potential biomarker for gastric cancer diagnosis and may be a novel therapeutic target because it inhibits the proliferation of gastric cancer cells [80]. This implies that tRF-33-P4R8YP9LON4VDP can be used for the diagnosis and treatment of gastric cancer in future. SVM prediction model for three tsRNAs, tRF-16L85J3KE, tRF-16-PSQP4PE, and tRF-21-RK9P4P9L0, can all predict lung adenocarcinoma [81]. tiRNA-1:33-GlyGCC1, tRF-1:32GlyGCC1, and tRF+1:T20SerTGA-1 may participate in the pathophysiological process of muscle-invasive bladder cancer [82]. Meanwhile, there are differences in the expression of tRF-1-32-chrm between glioblastoma and low-grade glioma tumor tissues [83]. tRFTyr increases lactate accumulation and promotes tumor progression in laryngeal squamous cell carcinoma, which can help develop new diagnostic biomarkers [84]. The levels of extracellular vesicles tRFLeuTAA005, tRFAsnGTT010, tRFAlaAGC036, tRFLysCTT049, and tRFTrpCA057 are significantly downregulated in non-small cell lung cancer, and may be promising biomarkers for the diagnosis [85]. Wang et al. [86] discovered six 5′-end-derived tRFs from tRNA, such as tRFGluCTC003, tRFGlyCCC007, tRFGlyCCC008, tRFLeuCAA003, tRFSerTGA001, and tRFSerTGA002. They may serve as potential novel biomarkers for early-stage breast cancer diagnosis. In addition, tiRNA-1:34GluTTC2 and tRF-60:76ArgACG1M2 may be associated with therapeutic strategies against multiple bone diseases [87]. Three tRF subtypes (tRF-3022b, tRF-3030b, and tRF-5008b) displayed elevated expression in colorectal cancer tissues compared to adjacent normal tissues, and their levels were also increased in plasma exosomes from patients when compared to healthy controls [88].
tsRNAs from patients with various tumors
| Disease | Sample types | tsRNA | Reference |
|---|---|---|---|
| Pancreatic cancer | Serum | tRFProAGG004,tRFLeuCAG002 | [81] |
| Pancreatic ductal adenocarcinoma | Plasma, Tissue | tsRNAValTAC41、tsRNAMetCAT37 | [82] |
| Chronic lymphocytic leukemia | CD5+/CD19+B cell | tRNAHis | [83] |
| Chronic lymphocytic leukemia, lung cancer | Tissue | tsRNA-46 and tsRNA-47 | [84] |
| Ovarian cancer | Tissue | tsRNA-29 | [84] |
| Hepatocellular carcinoma | Serum | tRFGlnTTG006 | [85] |
| Liver cancer | Plasma exosomal | tRNA-5 | [86] |
| Gastric cancer | Plasma | tRF-33-P4R8YP9LON4VDP | [87] |
| Lung adenocarcinoma | Plasma, Tissue | tRF-16-L85J3KE,tRF-21-RK9P4P9L0,tRF-16-PSQP4PE | [88] |
| Muscle-invasive bladder cancer | Tissue | tiRNA-1:33GlyGCC1,tRF-1:32GlyGCC1,tRF-+1:T20-SerTGA1 | [89] |
| Glioblastoma, Low-grade glioma | Tissue | tRF-1- 32-chrM | [90] |
| Laryngeal squamous cell carcinoma | Tissue | tRFTyr | [91] |
| Non-small cell lung cancer | Plasma exosomal | tRFLeuTAA005,tRFAsnGTT010,tRFAlaAGC036,tRFLysCTT049,tRFTrpCCA057 | [92] |
| Breast cancer | Plasma, Tissue | tRFGluCTC003,tRFGlyCCC007,tRFGlyCCC008, tRFLeuCAA003,tRFSerTGA001,tRFSerTGA002 | [93] |
| Multiple myeloma | Bone marrow | tiRNA-1:34GluTTC-2,tRF-60:76ArgACG1M2 | [94] |
| Colorectal cancer (CRC) | Plasma exosomal, Tissue | tRF-3022b,tRF-3030b,tRF-5008b | [95] |
2.2 tsRNA predicts therapeutic prognosis for tumor patients
In situ hybridization (ISH) scores for tRFProAGG004 and tRFLeuCAG002 may serve as valuable biomarkers for predicting postoperative survival time in patients with pancreatic cancer [74]. Balatti et al. discovered that ts-36 may play an important role in the malignant transformation of colon cells [76]. Additionally, high i-tRFGlyGCC is associated with shorter disease-free survival (DFS) in patients with colorectal cancer, and it is also associated with an increased likelihood of poorer overall survival in this group [89]. tRF-21-RKP4P9L0 is linked to the prognosis of lung adenocarcinoma (LUAD) [81]. In a therapeutic context, the inhibition of exosomal tRF-16-K8J7K1B enhances sensitivity to tamoxifen in breast cancer, suggesting that tRF-16-K8J7K1B could be a novel therapeutic target for overcoming tamoxifen resistance [90].
3. tsRNAs in patients with non-tumor diseases and clinical significance
Some distinct tsRNAs may act as novel diagnostic biomarkers for SLE, osteoporosis, AD, osteoporosis, and OSAHS. tRFHisGTG1 combined with anti-dsDNA was verified to be served as a biomarker for diagnosing systemic lupus erythematosus with an area under the curve of 0.95 (95% CI = 0.92-0.99), a sensitivity of 83.72%, and a specificity of 94.19% [91]. Compared with healthy individuals, tsRNA-10277 is significantly downregulated in the plasma exosomes from patients with steroid-induced osteonecrosis [92]. In idiopathic pulmonary arterial hypertension (PAH) [35], four upregulated tsRNAs were identified, namely i-tRF-31:54ValCa1, 5'tiRNA-31GluCTC16, tRF-3aAspGTC9, and tRF-3bTyrGTA4, and four downregulated tsRNAs were discovered, namely 5'tiRNA-33LysTTT4, i-tRF-8:32ValAAC2, i-tRF-2:30HisGTG-1, and i-tRF-15:31LysCTT1. Plasma tRF-25, tRF-38, and tRF-18 exosomes may serve as diagnostic biomarkers for the detection of osteoporosis [93]. La Ferlita et al. reported the tRF spectrum of atopic dermatitis (AD) and suggested that plasma exosome tRF-28-QSZ34KRQ590K may be a potential biomarker for pediatric patients with AD [94]. The expression levels of tRF-16-79MP9PD and tRF-28-OB1690PQR304 in the plasma of the children with obstructive sleep apnea-hypopnea syndrome (OSAHS) are significantly decreased, and may become new biomarkers for OSAHS diagnosis [95]. In patients with diabetes nephropathy, the expressions of tRF-5GluCTC, tRF-5AlaCGC, and tRF-5ValCAC are significantly upregulated, while the expressions of tRF-5GlyCCC, tRF-3GlyGCC, and tRF-3IleAAT are significantly downregulated [96]. tRF-36-F900BY4D84KRIME, tRF-23-87R8WP9IY, and tRF-40-86J8WPMN1E8Y7Z2R may play a crucial role in the occurrence and development of varicose veins [97].
4. Conclusion and Perspective
Although tRFs and tiRNAs have been paid attention to, it is the tip of the “iceberg”. Currently, tRNA sequencing is considered more challenging than mRNA sequencing. Single-cell tRNA-seq may reveal unique expression and/or modification patterns that reflect the regulatory mechanisms of controlling individual cell protein synthesis [98]. Clarifying the mechanism by which tsRNAs in single cells contribute to human diseases will provide profound insights into disease therapies.
tsRNA applications in therapeutic and prognosis of tumor patients
| Disease | Sample type | tsRNA | Reference |
|---|---|---|---|
| Pancreatic cancer | Tissue | tRFProAGG004,tRFLeuCAG002 | [81] |
| Colorectal cancer | Tissue | tsRNA-40 | [84] |
| Colorectal cancer | Tissue | i-tRFGlyGCC | [96] |
| Lung adenocarcinoma | Plasma, Tissue | tRF-21-RKP4P9L0 | [88] |
| Breast cancer | Serum | tRF-16-K8J7K1B | [97] |
tsRNAs from non-tumor diseases
| Disease | Sample types | tsRNAs | Reference |
|---|---|---|---|
| Systemic lupus erythematosus (SLE) | Serum | tRFHisGTG1 | [74] |
| Hormone Osteonecrosis of Femoral Head (SONFH) | Plasma exosome | tsRNA-10277 | [75] |
| Pulmonary arterial hypertension (PAH) | Plasma | i-tRF-31:54ValCAC1,5'tiRNA-31GluCTC16,tRF-3aAspGTC9,tRF-3bTyrGTA4,5'tiRNA-33LysTTT4,i-tRF-8:32ValAAC2,i-tRF-2:30HisGTG-1, i-tRF-15:31LysCTT1 | [37] |
| Osteoporosis | Plasma exosome | tRF-25, tRF-38, tRF-18 | [76] |
| Atopic dermatitis (AD) | Plasma exosome | tRF-28-QSZ34KRQ590K | [77] |
| Obstructive sleep apnea-hypopnea syndrome (OSAHS) | Plasma | tRF-16-79MP9PD, tRF-28-OB1690PQR304 | [78] |
| Diabetic nephropathy (DN) | Serum | tRF-5GluCTC, tRF-5AlaCGC,tRF-5ValCAC,tRF-5GlyCCC,tRF-3GlyGCC,tRF-3IleAAT | [79] |
| Varicose veins (VVs) | Vascular tissue | tRF-36-F900BY4D84KRIME,tRF-23-87R8WP9IY, tRF-40-86J8WPMN1E8Y7Z2R | [80] |
Although traditional miRNA sequencing is limited by the highly modified and complex structure of tRNAs, methodological innovations in recent years have significantly improved this situation. (1) AlkB-facilitated RNA methylation sequencing (ARM-seq), which used AlkB demethylase to remove m1A and m3C modifications, exerts efficient and quantitative sequencing to whole transcriptome tRNA, and solves the problem of reverse transcriptase truncation at modification sites [99]. (2) PANDORA-sequencing (PANDORA-seq) combines AlkB processing and T4 polynucleotide kinase (T4PNK)-mediated end repair to efficiently identify and sequence tsRNA with 2',3'-cyclic phosphate (2',3'-cP) termini [100]. This technique reveals large amounts of tsRNA and rsRNA that are previously missed by traditional methods and identifies key regulatory molecules in reproductive biology and cancer. (3) Although PANDORA-seq integrated repair of cP termini, earlier studies and specific optimization protocols (sometimes referred to as cP-seq strategies) had emphasized the importance of specific enrichment of tRNA halves produced by Angiogenin cleavage [101]. These methods convert unligable cP termini into standard linker ligation substrates by enzymatic conversion, and are key to resolving tsRNA dynamics in stress response. (4) In addition, YAMAT-sequence [102] provides an efficient alternative to detect tRNAs by optimizing linker ligation conditions without enzyme pretreatment. Meanwhile, sequencing methods capable of single-base resolution localization modification (including MLC-sequencing, other cutting-edge attempts, and validation strategies combined with mass spectrometry) are constantly being developed to further explore the specific impact of modification on tsRNA function. These technological advances suggest that tRNA/tsRNA sequencing is no longer an “insurmountable” challenge, but a new era of precision and panorama. Ignoring these advances would not accurately reflect the current state of scientific research.
Furthermore, to clarify the structural characteristics of tsRNAs, future studies should focus on elucidating the functions of their components. However, methodologies for identifying tsRNAs remain underdeveloped, necessitating the establishment of effective research approaches to systematically investigate tsRNAs structures and their mechanisms of action. Accumulating evidence suggests that tRFs are involved in multiple aspects of cancer and may serve as potential biomarkers for cancer diagnosis and prognosis. Currently, tsRNA nomenclature, such as tsRNA, tiRNA, sitRNA, semi tRNA, is still irregular. Many studies believe that tsRNA is a more appropriate name. Different sequencing methods yield different results, resulting in significant heterogeneity. Consistent quality control standards and standardized operations can reduce differences at certain extent. tsRNAs are unstable and easily degraded, which makes their detection difficulty. However, there are practical challenges to their clinical application. It needs to develop novel methods to overcome RNA degradation. In summary, as technology advances, an increasing number of tsRNAs will be identified and characterized in the future, and understanding their biological roles and clarifying their regulatory mechanisms is the most important and difficult task.
Acknowledgements
We thank Dr. Lin Chen and Yao Wu in Hunan University of Chinese Medicine for constructive suggestion.
Data accessibility statement
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.
Author contributions
Tang CC and Tang Q prepared the figures, the tables, and wrote the manuscript. Tang S and Tang F supervised the project, and reviewed and revised the manuscript. All authors read and approved the final manuscript.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Kirchner S, Ignatova Z. Emerging roles of tRNA in adaptive translation, signalling dynamics and disease. Nat Rev Genet. 2015;16:98-112
2. Mao M, Chen W, Huang X. et al. Role of tRNA-derived small RNAs(tsRNAs) in the diagnosis and treatment of malignant tumours. Cell Commun Signal. 2023;21:178
3. Phizicky EM, Hopper AK. tRNA biology charges to the front. Genes Dev. 2010;24:1832-60
4. Frank DN, Pace NR. Ribonuclease P: unity and diversity in a tRNA processing ribozyme. Annu Rev Biochem. 1998;67:153-80
5. Dubrovsky EB, Dubrovskaya VA, Levinger L. et al. Drosophila RNase Z processes mitochondrial and nuclear pre-tRNA 3' ends in vivo. Nucleic Acids Res. 2004;32:255-62
6. Maraia RJ, Lamichhane TN. 3' processing of eukaryotic precursor tRNAs. Wiley Interdiscip Rev RNA. 2011;2:362-75
7. Saad AAA, Zhang K, Deng Q. et al. The functions and modifications of tRNA-derived small RNAs in cancer biology. Cancer Metastasis Rev. 2025;44:38
8. Dong H, Ye C, Ye X. et al. The biological role and molecular mechanism of transfer RNA-derived small RNAs in tumor metastasis. Front Oncol. 2025;15:1560943
9. Wang C, Gao Y, Liu K. et al. tiRNA-HAR contributes to ischemic myocardial injury via facilitating HuR-mediated stability of p53. Transl Res. 2025;280:17-28
10. Li D, Zhang J, Han X. et al. Targeting tsRNA-1797 alleviates diabetes-induced vascular dysfunction through modulating purine metabolism. Diabetes. 2025;74:1247-61
11. Meng F, Bai H, Ke K. et al. tRF5-22-SerGCT-1 protects the heart against myocardial injury by targeting MSK1. Epigenomics. 2025;17:439-51
12. Zhang X, Shi J, Thakore P. et al. Mitochondrial small RNA alterations associated with increased lysosome activity in an Alzheimer's disease mouse model uncovered by PANDORA-seq. Int J Mol Sci. 2025;26:3019
13. Bai H, Meng F, Ke K. et al. The significance of small noncoding RNAs in the pathogenesis of cardiovascular diseases. Genes Dis. 2024;12:101342
14. Liu H, Hu W, Zhang L. et al. Plasma-derived exosomal i-tRF-LeuCAA as biomarker for Glioma diagnosis and promoter of epithelial-mesenchymal transition via TPM4 regulation. CNS Neurosci Ther. 2025;31:e70356
15. Winek K, Soreq H. Emerging roles of transfer RNA fragments in the CNS. Brain. 2025;148:2631-45
16. Liu M, Zhuang X, Zhang H. et al. tRNA-derived small RNAs in digestive tract diseases: Progress and perspectives. Genes Dis. 2024;12:101326
17. Rao J, Xia L, Li Q. et al. A 6-tsRNA signature for early detection, treatment response monitoring, and prognosis prediction in diffuse large B cell lymphoma. Blood Cancer J. 2025;15:79
18. Yang P, Li Z, Chen X. et al. Non-canonical small noncoding RNAs in the plasma extracellular vesicles as novel biomarkers in gastric cancer. J Hematol Oncol. 2025;18:39
19. Xu T, Hua H, Song F. et al. tRF-34-P4R8YP9LON4VHM promotes hepatocellular carcinoma progression and tumour cell-induced angiogenesis via the MEK/ERK pathway. J Cell Mol Med. 2025;29:e70560
20. Wu L, Chen X, Chen D. et al. A novel tRNA-derived fragment, tRF-20-M0NK5Y93 inhibits the malignant progression of non-small cell lung cancer by mediating PLOD1. Arch Biochem Biophys. 2025;770:110431
21. Hu H, Wu Y, Yuan S. et al. Identification of a novel tRNA-derived small RNA fragment, tRF-16-2YU04DE, with the potential of inhibiting endometrial cancer progression. Med Oncol. 2025;42:173
22. Li C, Zhong S, Chen J. et al. TsRNA-49-73-Glu-CTC: A promising serum biomarker in non-small cell lung cancer. PLoS One. 2025;20:e0320187
23. Lee YS, Shibata Y, Malhotra A. et al. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009;23:2639-49
24. Couvillion MT, Sachidanandam R, Collins K. A growth-essential tetrahymena Piwi protein carries tRNA fragment cargo. Genes Dev. 2010;24:2742-7
25. Thompson DM, Parker R. Stressing out over tRNA cleavage. Cell. 2009;138:215-9
26. Kumar P, Anaya J, Mudunuri SB. et al. Meta-analysis of tRNA derived RNA fragments reveals that they are evolutionarily conserved and associate with AGO proteins to recognize specific RNA targets. BMC Biology. 2016;14:78
27. Pliatsika V, Loher P, Magee R. et al. MINTbase v2.0: a comprehensive database for tRNA-derived fragments that includes nuclear and mitochondrial fragments from all The Cancer Genome Atlas projects. Nucleic Acids Research. 2018;46:D152-9
28. Schaefer M, Pollex T, Hanna K. et al. RNA methylation by Dnmt2 protects transfer RNAs against stress-induced cleavage. Genes Dev. 2010;24:1590-5
29. Cole C, Sobala A, Lu C. et al. Filtering of deep sequencing data reveals the existence of abundant dicer-dependent small RNAs derived from tRNAs. RNA. 2009;15:2147-60
30. Xie Y, Yao L, Yu X. et al. Action mechanisms and research methods of tRNA-derived small RNAs. Signal Transduct Target Ther. 2020;5:109
31. Kumar P, Anaya J, Mudunuri SB. et al. Meta-analysis of tRNA derived RNA fragments reveals that they are evolutionarily conserved and associate with AGO proteins to recognize specific RNA targets. BMC Biol. 2014;12:78
32. Kumar P, Mudunuri SB, Anaya J. et al. tRFdb: a database for transfer RNA fragments. Nucleic Acids Res. 2015;43:D141-5
33. Liao JY, Ma LM, Guo YH. et al. Deep sequencing of human nuclear and cytoplasmic small RNAs reveals an unexpectedly complex subcellular distribution of miRNAs and tRNA 3' trailers. PLoS One. 2010;5:e10563
34. Pereira M, Ribeiro DR, Pinheiro MM. et al. m5U54 tRNA hypomodification by lack of TRMT2A drives the generation of tRNA-derived small RNAs. Int J Mol Sci. 2021;22:2941
35. Guzzi N, Cieśla M, Ngoc PCT. et al. Pseudouridylation of tRNA-derived fragments steers translational control in stem cells. Cell. 2018;173:1204-16
36. Lee S, Kim J, Valdmanis PN. et al. Emerging roles of tRNA-derived small RNAs in cancer biology. Exp Mol Med. 2023;55:1293-304
37. Chen Y, Tang Y, Hou S. et al. Differential expression spectrum and targeted gene prediction of tRNA-derived small RNAs in idiopathic pulmonary arterial hypertension. Front Mol Biosci. 2023;10:1204740
38. Zhang X, He X, Liu C. et al. IL-4 Inhibits the biogenesis of an epigenetically suppressive PIWI-interacting RNA to upregulate CD1a molecules on monocytes/dendritic cells. J Immunol. 2016;196:1591-603
39. Chen Q, Yan M, Cao Z. et al. Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder. Science. 2016;351:397-400
40. Goodarzi H, Liu X, Nguyen HC. et al. Endogenous tRNA-derived fragments suppress breast cancer progression via YBX1 displacement. Cell. 2015;161:790-802
41. Krishna S, Yim DG, Lakshmanan V. et al. Dynamic expression of tRNA-derived small RNAs define cellular states. EMBO Rep. 2019;20:e47789
42. Li Z, Ender C, Meister G. et al. Extensive terminal and asymmetric processing of small RNAs from rRNAs, snoRNAs, snRNAs, and tRNAs. Nucleic Acids Res. 2012;40:6787-99
43. Maute RL, Schneider C, Sumazin P. et al. tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma. Proc Natl Acad Sci USA. 2013;110:1404-9
44. Karaiskos S, Naqvi AS, Swanson KE. et al. Age-driven modulation of tRNA-derived fragments in Drosophila and their potential targets. Biol Direct. 2015;10:51
45. Luo S, He F, Luo J. et al. Drosophila tsRNAs preferentially suppress general translation machinery via antisense pairing and participate in cellular starvation response. Nucleic Acids Res. 2018;46:5250-68
46. Gebetsberger J, Wyss L, Mleczko AM. et al. A tRNA-derived fragment competes with mRNA for ribosome binding and regulates translation during stress. RNA Biol. 2017;14:1364-73
47. Lalande S, Merret R, Salinas-Giegé T. et al. Arabidopsis tRNA-derived fragments as potential modulators of translation. RNA Biol. 2020;17:1137-48
48. Ivanov P, Emara MM, Villen J. et al. Angiogenin-induced tRNA fragments inhibit translation initiation. Mol Cell. 2011;43:613-23
49. Lyons SM, Kharel P, Akiyama Y. et al. eIF4G has intrinsic G-quadruplex binding activity that is required for tiRNA function. Nucleic Acids Res. 2020;48:6223-33
50. Blanco S, Dietmann S, Flores JV. et al. Aberrant methylation of tRNAs links cellular stress to neuro-developmental disorders. EMBO J. 2014;33:2020-39
51. Wang Q, Lee I, Ren J. et al. Identification and functional characterization of tRNA-derived RNA fragments (tRFs) in respiratory syncytial virus infection. Mol Ther. 2013;21:368-79
52. Keam SP, Sobala A, Ten Have S. et al. tRNA-derived RNA fragments associate with human multisynthetase complex (MSC) and modulate ribosomal protein translation. J Proteome Res. 2017;16:413-420
53. Cho H, Lee W, Kim GW. et al. Regulation of La/SSB-dependent viral gene expression by pre-tRNA 3' trailer-derived tRNA fragments. Nucleic Acids Res. 2019;47:9888-901
54. Yeung ML, Bennasser Y, Watashi K. et al. Pyrosequencing of small non-coding RNAs in HIV-1 infected cells: evidence for the processing of a viral-cellular double-stranded RNA hybrid. Nucleic Acids Res. 2009;37:6575-86
55. Ruggero K, Guffanti A, Corradin A. et al. Small noncoding RNAs in cells transformed by human T-cell leukemia virus type 1: a role for a tRNA fragment as a primer for reverse transcriptase. J Virol. 2014;88:3612-22
56. Sharma U, Conine CC, Shea JM. et al. Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals. Science. 2016;351:391-6
57. Zhou J, Liu S, Chen Y. et al. Identification of two novel functional tRNA-derived fragments induced in response to respiratory syncytial virus infection. J Gen Virol. 2017;98:1600-10
58. Deng J, Ptashkin RN, Chen Y. et al. Respiratory syncytial virus utilizes a tRNA fragment to suppress antiviral responses through a novel targeting mechanism. Mol Ther. 2015;23:1622-9
59. Han L, Lai H, Yang Y. et al. A 5'-tRNA halve, tiRNA-Gly promotes cell proliferation and migration via binding to RBM17 and inducing alternative splicing in papillary thyroid cancer. J Exp Clin Cancer Res. 2021;40:222
60. Zhou J, Wan F, Wang Y. et al. Small RNA sequencing reveals a novel tsRNA-26576 mediating tumorigenesis of breast cancer. Cancer Manag Res. 2019;11:3945-56
61. Cui H, Li H, Wu H. et al. A novel 3'tRNA-derived fragment tRF-Val promotes proliferation and inhibits apoptosis by targeting EEF1A1 in gastric cancer. Cell Death Dis. 2022;13:471
62. Kim HK, Xu J, Chu K. et al. A tRNA-derived small RNA regulates ribosomal protein S28 protein levels after translation initiation in humans and mice. Cell Rep. 2019;29:3816-24
63. Saikia M, Jobava R, Parisien M. et al. Angiogenin-cleaved tRNA halves interact with cytochrome C, protecting cells from apoptosis during osmotic stress. Mol Cell Biol. 2014;34:2450-63
64. Veneziano D, Di Bella S, Nigita G. et al. Noncoding RNA: Current deep sequencing data analysis approaches and challenges. Hum Mutat. 2016;37:1283-98
65. Anderson P, Ivanov P. tRNA fragments in human health and disease. FEBS Lett. 2014;588:4297-304
66. Shakeri R, Hosseinkhani S, Los MJ. et al. Role of the salt bridge between glutamate 546 and arginine 907 in preservation of autoinhibited form of Apaf-1. Int J Biol Macromol. 2015;81:370-4
67. Cosentino C, Toivonen S, Diaz Villamil E. et al. Pancreatic β-cell tRNA hypomethylation and fragmentation link TRMT10A deficiency with diabetes. Nucleic Acids Res. 2018;46:10302-18
68. Sarker G, Sun W, Rosenkranz D. et al. Maternal overnutrition programs hedonic and metabolic phenotypes across generations through sperm tsRNAs. Proc Natl Acad Sci U S A. 2019;116:10547-10556
69. Li X, Hernandez R, Zhang X. et al. A cholesterol-responsive hepatic tRNA-derived small RNA regulates cholesterol homeostasis and atherosclerosis development. Nat Commun. 2025;16:11043
70. Lei Y, Gan M, Wang K. et al. A novel tRF-Gly is associated with obesity development through post-transcriptional regulation of lipid metabolism. Cell Mol Biol Lett. 2025;31:6
71. Yang S, Li J, Mao S. et al. DNMT2-m5C-ACLY axis promotes lenvatinib resistance in hepatocellular carcinoma through histone acetylation-mediated notch pathway. Adv Sci (Weinh). 2026;13:e15931
72. Isacson S, Karlsson K, Zalavary S. et al. Small RNA in sperm-Paternal contributions to human embryo development. Nat Commun. 2025;16:6571
73. La Ferlita A, Alaimo S, Veneziano D. et al. Identification of tRNA-derived ncRNAs in TCGA and NCI-60 panel cell lines and development of the public database tRF explorer. Database (Oxford). 2019;2019:baz115
74. Jin F, Yang L, Wang W. et al. A novel class of tsRNA signatures as biomarkers for diagnosis and prognosis of pancreatic cancer. Mol Cancer. 2021;20:95
75. Xue M, Shi M, Xie J. et al. Serum tRNA-derived small RNAs as potential novel diagnostic biomarkers for pancreatic ductal adenocarcinoma. Am J Cancer Res. 2021;11:837-48
76. Veneziano D, Tomasello L, Balatti V. et al. Dysregulation of different classes of tRNA fragments in chronic lymphocytic leukemia. Proc Natl Acad Sci USA. 2019;116:24252-8
77. Balatti V, Nigita G, Veneziano D. et al. tsRNA signatures in cancer. Proc Natl Acad Sci USA. 2017;114:8071-6
78. Zhan S, Yang P, Zhou S. et al. Serum mitochondrial tsRNA serves as a novel biomarker for hepatocarcinoma diagnosis. Front Med. 2022;16:216-26
79. Zhu L, Li J, Gong Y. et al. Exosomal tRNA-derived small RNA as a promising biomarker for cancer diagnosis. Mol Cancer. 2019;18:74
80. Shen Y, Yu X, Ruan Y. et al. Global profile of tRNA-derived small RNAs in gastric cancer patient plasma and identification of tRF-33-P4R8YP9LON4VDP as a new tumor suppressor. Int J Med Sci. 2021;18:1570-9
81. Wang J, Liu X, Cui W. et al. Plasma tRNA-derived small RNAs signature as a predictive and prognostic biomarker in lung adenocarcinoma. Cancer Cell Int. 2022;22(1):59
82. Qin C, Chen ZH, Cao R. et al. Differential expression profiles and bioinformatics analysis of tRNA-derived small RNAs in muscle-invasive bladder cancer in a Chinese population. Genes (Basel). 2022;13:601
83. Tu M, Zuo Z, Chen C. et al. Transfer RNA-derived small RNAs (tsRNAs) sequencing revealed a differential expression landscape of tsRNAs between glioblastoma and low-grade glioma. Gene. 2023;855:147114
84. Zhao R, Yang Z, Zhao B. et al. A novel tyrosine tRNA-derived fragment, tRFTyr, induces oncogenesis and lactate accumulation in LSCC by interacting with LDHA. Cell Mol Biol Lett. 2023;28:49
85. Zheng B, Song X, Wang L. et al. Plasma exosomal tRNA-derived fragments as diagnostic biomarkers in non-small cell lung cancer. Front Oncol. 2022;12:1037523
86. Wang J, Ma G, Li M. et al. Plasma tRNA fragments derived from 5' ends as novel diagnostic biomarkers for early-stage breast cancer. Mol Ther Nucleic Acids. 2020;21:954-64
87. Xu C, Liang T, Zhang F. et al. tRNA-derived fragments as novel potential biomarkers for relapsed/refractory multiple myeloma. BMC Bioinformatics. 2021;22:238
88. Lu S, Wei X, Tao L. et al. A novel tRNA-derived fragment tRF-3022b modulates cell apoptosis and M2 macrophage polarization via binding to cytokines in colorectal cancer. J Hematol Oncol. 2022;15:176
89. Christodoulou S, Katsaraki K, Vassiliu P. et al. High intratumoral i-tRF-GlyGCC expression predicts short-term relapse and poor overall survival of colorectal cancer patients, independent of the TNM stage. Biomedicines. 2023;11:1945
90. Sun C, Huang X, Li J. et al. Exosome-transmitted tRF-16-K8J7K1B promotes tamoxifen resistance by reducing drug-induced cell apoptosis in breast cancer. Cancers (Basel). 2023;15:899
91. Yang P, Zhang X, Chen S. et al. A novel serum tsRNA for diagnosis and prediction of nephritis in SLE. Front Immunol. 2021;12:735105
92. Fang S, He T, Jiang J. et al. Osteogenic effect of tsRNA-10277-loaded exosome derived from bone mesenchymal stem cells on steroid-induced osteonecrosis of the femoral head. Drug Des Devel Ther. 2020;14:4579-91
93. Zhang Y, Cai F, Liu J. et al. Transfer RNA-derived fragments as potential exosome tRNA-derived fragment biomarkers for osteoporosis. Int J Rheum Dis. 2018;21:1659-69
94. Meng L, Jiang L, Chen J. et al. Transfer RNA-derived fragment tRF-28-QSZ34KRQ590K in plasma exosomes may be a potential biomarker for atopic dermatitis in pediatric patients. Exp Ther Med. 2021;21:489
95. Wu J, Cai X, Lu Y. et al. Plasma tRF-16-79MP9PD and tRF-28-OB1690PQR304 as potential biomarkers for 4- to 7-year-old children with obstructive sleep apnea-hypopnea syndrome. Front Pediatr. 2023;11:1141348
96. Huang C, Ding L, Ji J. et al. Expression profiles and potential roles of serum tRNA-derived fragments in diabetic nephropathy. Exp Ther Med. 2023;26:311
97. Yu C, Wang X, Hong Y. et al. Expression profile of tRNA-derived fragments and their potential roles in human varicose veins. Mol Med Rep. 2019;20:3191-201
98. Pan T. Modifications and functional genomics of human transfer RNA. Cell Res. 2018;28:395-404
99. Zheng G, Qin Y, Clark WC. et al. Efficient and quantitative high-throughput tRNA sequencing. Nat Methods. 2015;12:835-7
100. Shi J, Zhang Y, Tan D. et al. PANDORA-seq expands the repertoire of regulatory small RNAs by overcoming RNA modifications. Nat Cell Biol. 2021;23:676
101. Jodar M, Selvaraju S, Sendler E. et al. Reproductive Medicine Network. The presence, role and clinical use of spermatozoal RNAs. Hum Reprod Update. 2013;19:604-24
102. Shigematsu M, Honda S, Loher P. et al. YAMAT-seq: an efficient method for high-throughput sequencing of mature transfer RNAs. Nucleic Acids Res. 2017;45:e70
Author contact
![]() Corresponding author: Dr. Faqing Tang, Hunan Key Laboratory of Oncotarget Gene, Hunan Cancer Hospital, and The Affiliated Cancer Hospital of Xiangya School of Medicine, Central South University, Changsha 410013, China. Email: tangfqorg.cn.
Corresponding author: Dr. Faqing Tang, Hunan Key Laboratory of Oncotarget Gene, Hunan Cancer Hospital, and The Affiliated Cancer Hospital of Xiangya School of Medicine, Central South University, Changsha 410013, China. Email: tangfqorg.cn.