Impact Factor ISSN: 1837-9664

 Global reach, higher impact
Global reach, higher impactJ Cancer 2026; 17(6):1195-1205. doi:10.7150/jca.133243 This issue Cite
Research Paper
Assessing the Association Between Genetic Variants in ACE, SOD1, and PER3 and their Role in Breast Cancer Risk among Jordanian Women
Laith N. AL-Eitan1 ![]() , Fouad A. Almomani1, Mohammed S. Alorjani2, Maryam K. Alasmar1, Haneen O. Ali1, Mansour A. Alghamdi3,4
, Fouad A. Almomani1, Mohammed S. Alorjani2, Maryam K. Alasmar1, Haneen O. Ali1, Mansour A. Alghamdi3,4
1. Department of Biotechnology and Genetic Engineering, Jordan University of Science and Technology, Irbid 22110, Jordan.
2. Department of Pathology and Microbiology, College of Medicine, Jordan University of Science and Technology, Irbid 22110, Jordan.
3. Department of Anatomy, College of Medicine, King Khalid University, Abha 62529, Saudi Arabia.
4. Genomics and Personalized Medicine Unit, The Center for Medical and Health Research, King Khalid University, Abha 62529, Saudi Arabia.
Received 2026-2-16; Accepted 2026-5-30; Published 2026-6-10
Abstract

Background: Genetic and environmental factors regulate many physiological processes in the human body, and alterations in these processes may contribute to the development of various diseases, including breast cancer (BC), which is considered the most prevalent cancer among women and a leading cause of cancer-related mortality in the Jordanian population. Genes such as ACE, SOD1 and PER3 play important roles in regulating essential biological functions. These genes are involved in key physiological pathways, including blood pressure regulation, oxidative stress response and circadian rhythm maintenance, and genetic variants within them may influence susceptibility to cancer. Therefore, this study investigates the association between polymorphisms in the ACE, SOD1 and PER3 genes and the risk of breast cancer, with the aim of evaluating how these genetic variations relate to breast cancer susceptibility and clinical outcomes in Jordanian women.
Methods: Blood samples of 300 women diagnosed with breast cancer, along with 300 healthy participants, were collected, and DNA was extracted from them. Genetic variants in the ACE (rs1799752), SOD1 (rs36232792) and PER3 (rs57875989) genes are examined through employing direct PCR to amplify the target regions.
Results: The ACE (rs1799752) variant was observed to be associated with breast cancer susceptibility, with the I/I genotype increasing risk of breast cancer (OR = 5.138, 95% CI = 1.38-19.03, p = 0.014). No associations were observed for SOD1 (rs36232792) and PER3 (rs57875989) variants.
Conclusion: The rs1799752 polymorphism is suggested to have the potential of serving as a biomarker for breast cancer susceptibility in Jordanian women, as it is associated with elevating the risk.
Keywords: breast cancer, polymorphism, ACE, SOD1, PER3
Introduction
Breast cancer is the second leading cause of cancer-related deaths worldwide and is the most common cancer in women. It originates in breast cells, forming a tumor that can invade nearby tissues and metastasize to other parts of the body. Some alterations in breast cells can also lead to non-cancerous conditions, such as atypical hyperplasia, cysts, and benign tumors like intraductal papillomas [1]. Breast cancer often starts in the ducts or lobules of the breasts, which contain milk-producing cells organized into lobes and connected to the nipple through ducts [2]. Its development is influenced by genetic, hereditary, and various environmental risk factors. Treatment strategies depend on specific molecular features, including hormone receptor status (ER and PR), HER2 activation, gene mutations such as BRCA1/2 and PIK3CA, and immune markers such as TILs and PD-L1 [3].
There are different molecular subtypes of breast cancer, such as Luminal A, Luminal B, HER2-enriched, and basal-like. These subtypes are based on mRNA gene expression profiles. Each subtype has its own biological traits that help in improving patient outcomes by employing a personalized approach in treatment [4]. Several factors have been identified as significant risk factors for breast cancer, including genetic predisposition, family history, and reproductive factors such as age at menarche, parity and breastfeeding, as well as lifestyle-related factors including diet, obesity, smoking, alcohol consumption and exposure to radiation [5].
Angiotensin-converting enzyme (ACE) is an enzyme that controls several physiological processes in the cardiovascular system and is considered a dipeptidase enzyme, and its enzymatic activity is dependent on chloride and zinc ions, which are necessary for its action. While ACE exists in various forms, the primary isoform is a membrane-bound glycoprotein predominantly expressed in the lungs, anchored to the surfaces of cells that line blood vessels [6]. To maintain fluid and electrolyte balance, ACE supports homeostasis through its role in the renin angiotensin system (RAS). The RAS operates via two primary pathways: ACE/Ang II/AT1R, which promotes vascular constriction and cell proliferation and ACE2/Ang1-7/MasR, which generally has opposing effects. Dysregulation of the renin-angiotensin system (RAS), particularly the ACE/Ang II/AT1R pathway, has been linked to several cancers, including breast, ovarian and prostate cancers, and contributes to key processes such as cell proliferation, angiogenesis and metastasis [7, 8].
SOD1 (Cu/Zn superoxide dismutase) is an enzyme that breaks down superoxide radicals (O₂⁻) produced in the mitochondria and cytosol, helping to control oxidative stress in cells. What distinguishes SOD1 from other human superoxide dismutases is its high concentration in the cytosol. The other two forms are Mn superoxide dismutase (SOD2), located exclusively in the mitochondria and extracellular superoxide dismutase (SOD3), which also binds copper and zinc and is found in different cellular compartments, with SOD3 attached to the extracellular matrix. [9]. In addition to its well-established role as a superoxide dismutase, the SOD1 enzyme also exhibits peroxidase activity and possesses the capacity to bind DNA, suggesting additional roles in cellular processes [10]. The protein features an eight-stranded β-barrel structure with a catalytic copper ion, a structural zinc ion, an electrostatic loop and an intramolecular disulfide bond between cysteines 57 and 146. Variants of this gene specifically caused by missense mutations could affect the protein structure and function and are of significant interest for research studies of the role of SOD1 in different diseases [11].
PER3 is a major component of the circadian clock that works with proteins like PERIOD and CRYPTOCHROME (CRY) to regulate the body's internal rhythm by inhibiting core clock gene expression [12]. PER3 and PER1, PER2, CRY1, and CRY2 form complexes that move into the nucleus to suppress BMAL1 and CLOCK transcription factors. Disturbances in circadian rhythms are linked to higher cancer risk, with altered circadian gene expression being associated with cancer progression [13]. PER3 is in the 1p36 chromosomal region, frequently deleted in human cancers, particularly breast cancer [14, 15]. Its expression fluctuates in peripheral tissues and organs as a component of the negative branch of the core molecular circadian clock feedback loop [16, 17, 18]. Patients with breast and other cancers exhibit reduced PERIOD clock gene expression in their tumors compared to adjacent normal tissue [19, 20]. This and additional data suggest that PERIOD clock genes function as tumor suppressors [14, 21].
The association between polymorphisms in the ACE (rs1799752), SOD1 (rs36232792), and PER3 (rs57875989) genes and breast cancer risk yielded varied results in different populations. Additionally, the research examining the combined impact of these specific genetic variants in breast cancer patients within the Jordanian population is insufficient. In this regard, this study seeks to address the gap by examining the prevalence of these genetic polymorphisms among Jordanian women. Moreover, this research also aims to explore the potential impacts of these genetic differences on breast cancer susceptibility as well as the survival rates of breast cancer patients.
Methods
Subjects
A case-control study was designed to explore the link between genetic polymorphisms in the ACE, SOD1 and PER3 genes and the risk of developing breast cancer. This study included 600 participants, with equal numbers of 300 breast cancer patients and 300 healthy controls who were not related to each other. Participants were recruited randomly from the chemotherapy clinics at King Abdullah University Hospital (KAUH) and King Hussein Medical Center. The study protocol was approved by the Human Ethics Committee of Jordan University of Science and Technology and KAUH (No.: 9/143/2021). Participant's informed consent was obtained prior to sample collection. Comprehensive data for the patients, including clinical history, demographic data, diagnostic results, and medication data, was collected from the electronic medical records of the patients. For the patients to be included in the study, they had to have a confirmed diagnosis of breast cancer based on histopathological results. In addition, the patients had to be negative for HIV, HBV, and HCV infections. Furthermore, the patients had to have comprehensive data recorded in the KAUH patient registry. Exclusion criteria included patients who failed to give consent for the study, patients who had undergone blood transfusions during surgery, and patients whose clinical documentation was inadequate.
Candidate gene and polymorphisms selection
To select polymorphisms for this study, we focused on significant polymorphisms in the ACE, SOD1 and PER3 genes reported in breast cancer patients from diverse populations globally. The selected polymorphisms underwent further validation using various predictive online tools and specific web servers to assess their potential impact. These polymorphisms were compared and verified through several databases, including the NCBI SNP database (https://www.ncbi.nlm.nih.gov/snp/), HaploReg v4.2 (https://pubs.broadinstitute.org/mammals/haploreg/haploreg.php), Ensembl (http://www.ensembl.org/index.html), RegulomeDB (https://regulomedb.org/regulome-search/), rSNPBase 3.1 (http://rsnp3.psych.ac.cn/) and the SNP Function Prediction tool (https://snpinfo.niehs.nih.gov/snpinfo/snpfunc.html). This is to ensure that the polymorphisms used for the research were relevant and accurate.
The RegulomeDB database shows that the polymorphisms rs1799752 in the ACE gene, rs36232792 in SOD1 and rs57875989 in PER3 each received a score of 4, along with a rating of 0.60906. The results suggest that these polymorphisms could affect binding sites and contribute to the regulation of gene expression. Moreover, the HaploReg v4.2 database showed functional annotations of those genes, where the rs1799752 variant, located in the intronic region within the ACE gene, affects five different binding motifs. The rs36232792 in the SOD1 gene is a synonymous coding SNV that potentially modifies twelve motifs, impacting histone marks in promoters and enhancers and influencing transcription factor binding sites. Meanwhile, the rs57875989 variant of the PER3 gene alters the Zfp410 motif and could also influence histone marks in promoters and enhancers. These findings suggest that the polymorphisms in ACE, SOD1 and PER3 may indirectly contribute to breast cancer development by impacting the regulation of transcription factors associated with these genes.
DNA extraction
Using the Puregene® Blood Core Kit A (Qiagen), genomic DNA was isolated from blood samples by adhering to the manufacturer's protocol. The isolated DNA purity and concentration were assessed with a NanoDrop spectrophotometer and verified through gel electrophoresis.
Genotyping
Genotyping of the rs1799752 variant in the ACE gene, rs36232792 in SOD1 and rs57875989 in PER3 was conducted using direct PCR. Each PCR reaction was carried out in a total volume of 25 µL, which included nuclease-free water, a 2x master mix, 10 µM of both forward and reverse primers and template DNA. The specific primer sequences used for the current study have been previously reported [22-24]. Further details on the reaction conditions can be found in Table 1. Genotyping quality was further validated by repeating the genotyping for randomly selected samples and verifying genotype consistency. Samples with unclear or ambiguous results were re-analyzed to confirm genotype assignment.
Genotyping parameters and PCR product characteristics.
| Polymorphism | Genotyping | Annealing temperature | Allele | Fragment size (bp) |
|---|---|---|---|---|
| ACE (rs1799752) | Direct PCR | 62 | I D | 490 190 |
| SOD1 (rs36232792) | Direct PCR | 62 | I D | 297 247 |
| PER3 (rs57875989) | Direct PCR | 68 | 5R 4R | 257 193 |
Statistical analyses
Statistical analyses were conducted using various methods and tools to ensure reliable and accurate results. To assess Hardy-Weinberg equilibrium, allele and genotype frequencies, inheritance models and the relationship between haplotypes and disease status, the SNPstat tool (https://www.snpstats.net/start.htm) was utilized. In addition, odds ratios (OR) and 95% confidence intervals (CI) were determined, with statistical significance defined by p-values below 0.05. The Statistical Package for the Social Sciences (SPSS) software, version 26.0 (SPSS, Inc., Chicago, IL), was used to evaluate genotype-phenotype correlations using Pearson's chi-square test and one-way ANOVA. To account for rare genotypes with small sample counts, Fisher's exact test was applied when expected counts were less than five, while the chi-square test was used for comparisons with sufficient counts. Post hoc statistical power was estimated using G*Power (v3.1.9.7), utilizing the observed genotype frequencies in cases and controls, with a two-tailed test for comparisons between independent groups and an alpha level of 0.05. Multivariable binary logistic regression analysis was conducted to determine whether the studied polymorphisms were independent risk factors for disease susceptibility after adjustment for potential confounding variables. Survival analysis was performed using the Kaplan-Meier method in R (version 4.5.3), and differences between groups were assessed using the log-rank test. To adjust for multiple testing, the appropriate number of polymorphisms was calculated according to a method described in a prior study [25]. The significance level was further refined using the Bonferroni correction, where it was set to α/n, with α = 0.05 and n representing the total number of tests performed [26].
Results
Patient characteristics
ACE, SOD1, and PER3 gene polymorphisms were utilized in this case-control study to explore their association with the risk of developing breast cancer. 300 unrelated female patients with breast cancer and 300 unrelated healthy female controls from the Jordanian population were included in the study. The mean age of the breast cancer patients was 52.32 ± 11.39 years, with a median age of 51 years and an age range of 25-85 years. Moreover, 160 patients reported a family history of cancer, representing 56.34% of the breast cancer group.
Hardy-Weinberg Equilibrium (HWE) and minor allele frequencies
The genetic variations rs1799752, rs36232792, and rs57875989 came under a thorough scrutiny of the minor allele frequencies in breast cancer patients and in normal persons, which was fundamental in defining the possible variations in the distribution of genetic variations between both categories, suggesting a possible link between them and the risk of breast cancer. The HWE test was applied to confirm the genetic information since it determines whether the data on the allele and genotype frequencies within a population fit expectations from random mating and is a fundamental marker of the genetic constitution of the investigated population.
For most of the polymorphisms analyzed, the genotype frequencies in both cases and controls were in consistent with Hardy-Weinberg equilibrium, indicating that the study populations were genetically stable and unlikely to be affected by factors such as selection, mutation, or genotyping errors that could alter allele distribution. However, the rs1799752 polymorphism in the ACE gene showed a clear deviation from Hardy-Weinberg equilibrium in both groups. Detailed data supporting these findings are presented in Table 2.
Hardy-Weinberg equilibrium values and minor allele frequencies of ACE, SOD1 and PER3 polymorphisms.
| Gene | Control (n=300) | Cases (n = 300) | ||||
|---|---|---|---|---|---|---|
| MA | MAF | HWE p-value | MA | MAF | HWE p-value | |
| ACE (rs1799752) | I | 30% | 0.0053 | I | 27% | < 0.0001 |
| SOD1 (rs36232792) | D | 15% | 0.37 | D | 12% | 0.062 |
| PER3 (rs57875989) | 5R | 46% | 0.16 | 5R | 46% | 0.25 |
MA: minor allele.
MAF: minor allele frequency.
HWE: Hardy-Weinberg equilibrium
Allele and genotype distributions and their association with breast cancer risk
Detailed information on allele frequencies and genotype distributions, along with unadjusted and adjusted odds ratios obtained using multivariable logistic regression analysis controlling for age, sex, BMI, and smoking status, is presented in Table 3. Associations of genotypes were evaluated under a codominant genetic model. For the ACE rs1799752 variant, the I/I genotype was associated with an increased risk of breast cancer in both unadjusted analysis (OR = 4.12, 95% CI: 1.34-12.62, p = 0.009) and after adjustment (adjusted OR = 5.14, 95% CI: 1.38-19.03, p = 0.014). While the SOD1 rs36232792 variant, the D/D genotype, showed a significant association in the unadjusted analysis, this association was attenuated and no longer statistically significant after adjustment (adjusted OR = 7.21, 95% CI: 0.88-59.12, p = 0.066). No significant associations were observed for the PER3 rs57875989 variant in either unadjusted or adjusted analyses.
Allele and genotype distributions with unadjusted and adjusted ORs for ACE, SOD1, and PER3 variants.
| Gene | SNP_ID | Allele/ Genotype | Control (n=300) | Cases (n=300) | Unadjusted OR (95% CI) | p-value | Adjusted OR (95% CI) | p-value |
|---|---|---|---|---|---|---|---|---|
| ACE | rs1799752 | D I | 422 (70%) 178 (30%) | 438 (73%) 162 (27%) | 1.00 1.14 (0.88-1.46) | 0.337 | --- | --- |
| D/D I/D I/I | 138 (46%) 146 (49%) 16 (5%) | 142 (47%) 154 (51%) 4 (1%) | 1.00 0.98 (0.70-1.36) 4.12 (1.34-12.62) | 0.934 0.009 | 1.00 1.04 (0.72-1.51) 5.14 (1.38-19.03) | 0.818 0.014 | ||
| SOD1 | rs36232792 | I D | 508 (85%) 92 (15%) | 525 (88%) 75 (12%) | 1.00 0.79 (0.57-1.10) | 0.182 | --- | --- |
| I/I I/D D/D | 217 (72%) 74 (25%) 9 (3%) | 226 (75%) 73 (24%) 1 (0%) | 1.00 1.06 (0.73-1.52) 9.37 (1.53-103.4) | 0.849 0.011 | 1.00 0.92 (0.60-1.39) 7.21 (0.88-59.12) | 0.685 0.066 | ||
| PER3 | rs57875989 | 4R 5R | 325 (54%) 275 (46%) | 324 (54%) 276 (46%) | 1.00 0.99 (0.79-1.27) | 0.999 | --- | --- |
| 4R/4R 4R/5R 5R/5R | 94 (31%) 137 (46%) 69 (23%) | 82 (27%) 160 (53%) 58 (19%) | 1.00 0.75 (0.52-1.09) 1.04 (0.65-1.66) | 0.129 0.907 | 1.00 0.13 (0.02-1.06) 0.14 (0.02-1.14) | 0.057 0.066 |
P-values < 0.0167 (0.05/# of variants, 0.05/3 = 0.0167 after applying multiple comparisons) are considered significant.
Analyses using different genetic models
The association between polymorphisms in the ACE, SOD1, and PER3 genes and breast cancer risk was investigated through various genetic models. Odds ratios (ORs) for these associations are detailed in Table 4. Significant associations were observed for the ACE (rs1799752) polymorphism under the recessive inheritance models (OR = 3.82, P = 0.011). The post hoc power for this association was 0.78. In contrast, no significant association was found for the SOD1 and PER3 polymorphisms across the genetic models analyzed.
Analyses of ACE, SOD1, and PER3 polymorphisms using different genetic models.
| Polymorphism | Model | Genotype | Cases (%) | Controls (%) | OR (95% CI) | P value |
|---|---|---|---|---|---|---|
| ACE (rs1799752) | Dominant | D/D I/D-I/I | 142 (47.3%) 158 (52.7%) | 138 (46%) 162 (54%) | 1.00 1.06 (0.77-1.45) | 0.74 |
| Recessive | D/D-I/D I/I | 296 (98.7%) 4 (1.3%) | 284 (94.7%) 16 (5.3%) | 1.00 3.82 (1.33-10.97) | 0.011 | |
| Overdominant | D/D-I/I I/D | 146 (48.7%) 154 (51.3%) | 154 (51.3%) 146 (48.7%) | 1.00 0.90 (0.65-1.24) | 0.51 | |
| SOD1 (rs36232792) | Dominant | I/I I/D-D/D | 226 (75.3%) 74 (24.7%) | 217 (72.3%) 83 (27.7%) | 1.00 1.17 (0.81-1.68) | 0.4 |
| Recessive | I/I-I/D D/D | 299 (99.7%) 1 (0.3%) | 291 (97%) 9 (3%) | 1.00 6.51 (1.15-36.69) | 0.021 | |
| Overdominant | I/I-D/D I/D | 227 (75.7%) 73 (24.3%) | 226 (75.3%) 74 (24.7%) | 1.00 1.02 (0.70-1.48) | 0.92 | |
| PER3 (rs57875989) | Dominant | 4R/4R 4R/5R-5R/5R | 82 (27.3%) 218 (72.7%) | 94 (31.3%) 206 (68.7%) | 1.00 0.82 (0.58-1.17) | 0.28 |
| Recessive | 4R/4R-4R/5R 5R/5R | 242 (80.7%) 58 (19.3%) | 231 (77%) 69 (23%) | 1.00 1.25 (0.84-1.85) | 0.27 | |
| Overdominant | 4R/4R-5R/5R 4R/5R | 140 (46.7%) 160 (53.3%) | 163 (54.3%) 137 (44.7%) | 1.00 0.74 (0.53-1.01) | 0.95 |
P-values < 0.0167 (0.05/# of variants, 0.05/3 = 0.0167 after applying multiple comparisons) are considered significant.
The clinical features and genotypic associations of polymorphisms
As outlined in Table 5, statistical analyses were performed to explore the association between ACE, SOD1, and PER3 polymorphisms and clinical outcomes in breast cancer patients. The results indicated that none of the PER3 (rs57875989), SOD1 (rs36232792), or PER3 (rs57875989) polymorphisms were associated with the clinical trait of breast cancer (p > 0.0167).
Association between ACE, SOD1, and PER3 polymorphisms and clinical features in breast cancer patients.
| Clinical Outcome | ACE | SOD1 | PER3 |
|---|---|---|---|
| Age | 0.374b 0.6883 | 1.8458b 0.160925 | 2.21639b 0.110814 |
| Age at BC Diagnosis | 0.04081b 0.96 | 1.40986b 0.245812 | 3.36033b 0.036067 |
| Stage of BC | 3.7051b 0.05727 | 1.59214b 0.208816 | 1.50217b 0.227998 |
| Age at First Pregnancy | 0.03022b 0.9702 | 2.95502b 0.054207 | 0.60699b 0.545978 |
| Body Mass Index | 2.0104b 0.1358 | 2.21565b 0.110895 | 1.1581b 0.315562 |
| Age of Menarche | 0.9335b 0.3945 | 2.88466b 0.057598 | 0.82813b 0.438019 |
| Age of Menopause | 0.08086b 0.9223 | 1.5338b 0.217803 | 2.72881b 0.067402 |
| Breastfeeding Status | 2.0784a 0.35373 | 0.755a 0.6856 | 1.305a 0.5207 |
| Family History of Cancer | 0.0492b 0.97572 | 4.007a 0.1349 | 1.016a 0.6017 |
| Other Types of Cancer | 4.6375a 0.098396 | 1.502a 0.4719 | 2.877a 0.2373 |
| Polycystic ovary syndrome (PCOS) | 0.355a 0.8374 | 0.78a 0.6771 | 0.9743a 0.614369 |
| Uterine Fibroid | 0.42a 0.8106 | 2.347a 0.3093 | 0.129a 0.9375 |
| Benign Breast Tumor | 0.086a 0.9579 | 1.489a 0.4750 | 2.4742a 0.290227 |
| Estrogen Receptor | 0.101a 0.9508 | 0.902a 0.6370 | 1.317a 0.5176 |
| Progesterone Receptor | 0.098a 0.9522 | 0.396a 0.8204 | 0.505a 0.7769 |
| Human Epidermal Growth Factor Receptor 2 (HER2) | 3.285a 0.1935 | 2.373a 0.3053 | 0.465a 0.7925 |
| Axillary Lymph Node Metastasis | 1.697a 0.4281 | 0.626a 0.7312 | 2.074a 0.3545 |
| Lymph vascular Invasion | 0.23a 0.6315 | 0.053a 0.9738 | 0.062a 0.9695 |
| Distant Metastasis | 0.145a 0.9301 | 1.775a 0.4117 | 1.911a 0.3846 |
| Allergy | 0.96a 0.6188 | 1.232a 0.5401 | 1.9413a o.378829 |
| Smoking | 3.173a 0.2046 | 0.802a 0.6697 | 0.113a 0.9451 |
A Parson's Chi-squared test was used to determine genotype-phenotype association (chi-square value).
b Analysis of variance (ANOVA) test was used to determine genotype-phenotype association (f-ratio value).
P-values < 0.0167 (0.05/# of variants, 0.05/3 = 0.0167 after applying multiple comparisons) are considered significant.
Logistic regression analysis
Multivariable binary logistic regression analysis was performed to evaluate whether the studied polymorphisms were independently associated with disease susceptibility after adjusting for potential confounding factors, including age, BMI, smoking status, age at menarche and family history of cancer. The results are presented in Supplementary Table S1.
For the ACE polymorphism, the I/I genotype was significantly associated with increased disease risk compared with the reference genotype (D/D) (OR = 5.14, 95% CI = 1.39-19.03, P = 0.014). In contrast, no significant associations were observed for genotypes of the SOD1 or PER3 polymorphisms (P > 0.05). However, family history of cancer was significantly associated with increased disease risk in all models (P < 0.001), while age, BMI, smoking status and age at menarche were not significant predictors.
Survival analysis of breast cancer patients
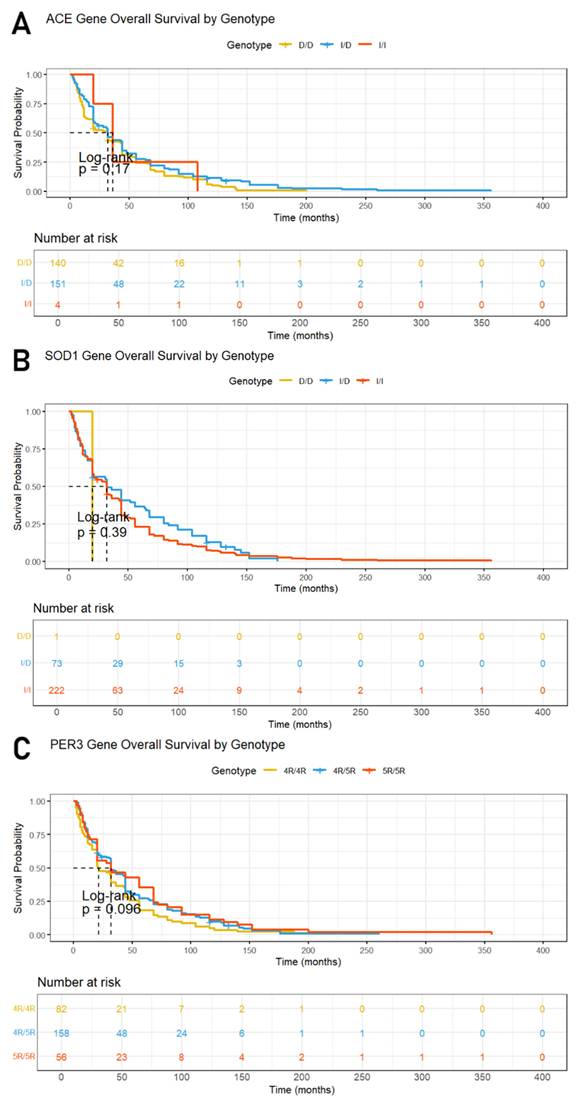
Kaplan-Meier survival curves and the log-rank test were employed to evaluate overall survival rates among breast cancer patients, as presented in Figure 1. The median follow-up time for the study cohort was three years, during which five deaths occurred. The results showed no significant differences in survival times related to the ACE (rs1799752), SOD1 (rs36232792) and PER3 (rs57875989) variants, as evidenced by p-values of 0.17, 0.39 and 0.096, respectively, indicating that these genetic polymorphisms do not significantly impact patient survival in this study.
Kaplan-Meier curves and the log-rank test for overall survival (OS) according to (A) ACE (rs1799752), (B) PER3 (rs57875989) and (C) SOD1 (rs36232792) gene polymorphism genotypes.
Discussion
Breast cancer is the most common cancer type among women around the world and presents a major public health problem because of its diversity. Genetic predisposition represents a major risk factor for its development and progression. The onset and progression of several cancers have been linked to alterations in ACE expression, with elevated ACE levels often associated with more aggressive disease and poorer clinical outcomes. Furthermore, activation of the ACE/Ang-II/AT1R signaling pathway promotes cellular proliferation, invasion, angiogenesis, epithelial-mesenchymal transition and therapy resistance in cancer models [27, 28].
The rs1799752 insertion/deletion (I/D) polymorphism in the ACE gene has been widely investigated for its potential involvement in breast cancer development. Studies have indicated that the D allele of rs1799752 is associated with significantly higher circulating levels of the enzyme encoded by ACE compared with the I allele [29]. This functional difference may influence angiotensin II production and consequently modulate pathways involved in tumor growth and metastasis. Based on this mechanism, Koh et al. proposed that women carrying the low-activity I allele may have reduced production of angiotensin II, which could potentially confer a lower susceptibility to breast cancer [30]. Therefore, the ACE I/D polymorphism may represent a potential biomarker for evaluating breast cancer risk and clinical outcomes [22, 31].
Our study has identified a relationship between breast cancer susceptibility and the rs1799752 variant in the ACE gene, which is in accordance with previous findings in different populations, including Egyptian [22], North Indian [32], Turkish [33] and Dutch cohorts [34], as well as meta-analyses conducted in Asian and Caucasian populations [31]. However, two other meta-analyses reported conflicting findings, indicating no significant association between the ACE I/D polymorphism and breast cancer risk [35, 36]. Similarly, no significant relationship between the ACE I/D polymorphism and breast cancer susceptibility was observed in a Pakistani population [37].
Superoxide dismutase 1 (SOD1) is a key enzyme involved in the regulation of reactive oxygen species (ROS) in breast cancer cells. While SOD2 primarily regulates ROS within the mitochondria and is often downregulated due to reduced SIRT3 activity—resulting in elevated mitochondrial ROS levels—cancer cells may compensate by overexpressing SOD1 in the cytoplasm and mitochondrial intermembrane space [38]. This compensatory mechanism helps limit excessive superoxide accumulation and maintain mitochondrial integrity. Inhibition of SOD1 has been shown to induce mitochondrial damage, highlighting its essential role in protecting cancer cells from oxidative stress and supporting their survival and proliferation [38].
Evidence suggests that SOD1 plays an important role in breast cancer development and tumor progression, independent of specific oncogenic drivers. Elevated SOD1 expression has been associated with poorer survival outcomes and increased metastatic potential. Although SOD1 appears to be essential for cancer cell growth, it is not required for normal cell proliferation [39]. The rs36232792 polymorphism, characterized by a 50-bp insertion/deletion in the promoter region of the SOD1 gene, may influence transcriptional regulation. Experimental in vitro studies have demonstrated that the 50-bp deletion reduces promoter activity and mRNA expression by eliminating two Sp1 transcription factor binding sites [40]. Consequently, carriers of the deletion allele may exhibit reduced SOD1 expression and impaired detoxification of reactive oxygen species (ROS), which could compromise genomic stability and contribute to interindividual differences in cancer susceptibility [21, 41].
Our study demonstrated that the rs36232792 polymorphism in the SOD1 gene is significantly associated with increased breast cancer risk. This finding is consistent with previous research in the Iranian population, which also reported no significant link between this variant and breast cancer susceptibility [42]. In contrast, studies in the Mexican population observed a significant association [43]. These discrepancies suggest that the impact of the rs36232792 polymorphism on breast cancer risk may be influenced by population-specific genetic backgrounds and environmental factors, emphasizing the importance of considering such differences in genetic association studies.
The Period Circadian Regulator 3 (PER3) gene have been implicated in breast cancer, with evidence suggesting that PER3 expression is reduced in tumor tissues. Lower PER3 levels have been associated with advanced tumor stage, higher metastasis risk and more aggressive cancer features, including ER negativity, higher histological grade and increased recurrence [14, 44]. PER3 appears to inhibit cancer progression by suppressing the MEK/ERK signaling pathway, which is crucial for cell proliferation and survival, highlighting its potential as a prognostic biomarker and therapeutic target [44]. Located on chromosome 1p36, a region frequently deleted in breast cancer, PER3 has also attracted attention due to its role in circadian regulation. Disrupted circadian rhythms and altered sleep patterns are linked to increased breast cancer risk, suggesting that circadian-regulating genes like PER3 may influence susceptibility [45]. Circadian genes, including PER3, can affect hormone secretion, further connecting circadian disruption with tumorigenesis [14].
The PER3 gene contains a variable number tandem repeat (VNTR) polymorphism (rs57875989) in exon 18, which consists of four or five copies of a 54-bp repeat sequence, which generates three genotypes: 4R/4R, 4R/5R, and 5R/5R. The 5-repeat allele introduces additional phosphorylation motifs within the PER3 protein, potentially altering phosphorylation-dependent regulation of circadian hormone secretion [46]. These functional changes have been linked to differences in sleep patterns and disorders, and carriers of the 5/5 genotype have been reported to exhibit increased breast cancer susceptibility, particularly in premenopausal women [47,48].
Our study did not detect a significant association between the PER3 rs57875989 polymorphism and breast cancer risk. This contrasts a previous study in Indian women, which reported an increased risk associated with the 5R genotype [46], while research in Caucasian women from Connecticut, USA, found that both heterozygous and homozygous carriers of the 5R allele exhibited elevated breast cancer risk, particularly among premenopausal individuals [49]. In line with our findings, a study in the Dutch population, together with a comprehensive meta-analysis, reported only a non-significant trend toward increased breast cancer risk among carriers of the 5-repeat allele, most pronounced in homozygous individuals [24].
In our study, Hardy-Weinberg equilibrium (HWE) tests were performed for all selected polymorphisms in both cases and controls. Most polymorphisms conformed to HWE expectations, except for rs1799752 of the ACE gene, which showed a significant deviation (p < 0.05). This deviation may be influenced by several factors, notably the relatively small sample size, which can amplify the effects of genetic drift and affect expected genotype distributions.
HWE tests evaluate whether genotype frequencies in a population conform to expectations under random mating and no evolutionary pressures. Deviations can result from mutation, selection, gene flow or non-random mating. In the present case, the reason behind the deviation of the findings of the present study could be attributed to the presence of consanguineous marriages, which are common among the Jordanian population. Such non-random mating reduces genetic diversity and increases the likelihood of inheriting identical alleles, potentially distorting genotype frequencies and disrupting HWE [50, 51].
Previous research has indicated that deviation of certain SNPs from Hardy-Weinberg equilibrium does not necessarily reflect genotyping error and may instead reflect a true association with disease risk; consequently, excluding such variants may lead to the omission of biologically relevant genetic effects [52-55]. Therefore, the basis of exclusion was not the violation of the Hardy-Weinberg equilibrium principle. The sensitivity analysis gave comparable results, validating the inclusion of this SNP in the accurate description of the population's genetic composition.
Statistical power analysis was conducted based on genotype frequencies under the most significant genetic models identified for each polymorphism. The ACE (rs1799752) variant showed significant associations with breast cancer susceptibility under both codominant and recessive models; however, the corresponding statistical power was 0.78, reflecting moderate power. Accordingly, these findings should be interpreted with caution and warrant validation in larger independent cohorts. These findings demonstrate the significance of incorporating power into the genotype-based inheritance model, as this would allow for a more accurate assessment of the genetic associations involved. However, the relatively low power exhibited by these findings indicates that additional research needs to be conducted on a larger scale in order to confirm these associations.
This study has several limitations that should be considered when interpreting the findings. Although the total sample size of 600 participants, including 300 cases and 300 controls, is comparable to many regional genetic association studies, it may still limit the statistical power to detect weak gene-disease associations. In particular, rare genotypes, such as the D/D genotype of the SOD1 and the I/I genotype of ACE, were infrequent, which may increase confidence interval width and reduce the precision of odds ratio estimates. Therefore, the possibility of statistical bias or reduced sensitivity in detecting modest genetic effects cannot be excluded. Therefore, larger studies with greater sample sizes are warranted to validate the present findings and further clarify the role of these polymorphisms in disease susceptibility.
Conclusion
The research provides essential insights into the relationship between ACE, SOD1, and PER3 polymorphisms and breast cancer risk in Jordan. Our findings indicate that the rs1799752 polymorphism in the ACE gene is significantly associated with breast cancer susceptibility in Jordanian women, suggesting its potential as a genetic marker for risk assessment. Although these findings could enhance patient stratification and risk assessment, further large-scale and functional studies are needed to confirm these associations. This study is pioneering in its attention to Jordanian Arab descendants, investigating the impact of genetic polymorphisms and ethnicity on breast cancer risk. It contributes valuable insights into personalized medicine for breast cancer by suggesting the need to study understudied ethnic groups. The research emphasizes the importance of expanding scientific exploration to understand disease risk factors and mechanisms comprehensively.
Supplementary Material
Supplementary table.
Acknowledgements
Funding
This research was funded by the Deanship of Research at Jordan University of Science and Technology, grant number RN: 20240119.
Ethics committee approval and patient consent
This study was performed in line with the principles of the Declaration of Helsinki. Approval was granted by the Institutional Review Board (IRB) committee at the Jordan University of Science and Technology (Date: 8 September 2021, No: 9/143/2021). Written informed consent was obtained from all subjects involved in the study.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Author contributions
Conceptualization, L.N.A.-E; methodology, L.N.A.-E and F.A.A.; validation, L.N.A.-E, M.S.A., and M.A.A.; formal analysis, L.N.A.-E, F.A.A., M.S.A, M.K.A., H.O.A., and M.A.A.; investigation, M.K.A. and H.O.A.; resources, M.S.A.; data curation, M.K.A. and H.O.A.; writing—original draft preparation, L.N.A.-E and F.A.A.; writing—review and editing, L.N.A.-E, F.A.A., M.S.A, M.K.A., H.O.A., and M.A.A.; supervision, L.N.A.-E and F.A.A.; project administration, L.N.A.-E.; funding acquisition, F.A.A. All authors have read and agreed to the published version of the manuscript.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Obeagu EI, Obeagu GU. Breast cancer: A review of risk factors and diagnosis. Medicine (Baltimore). 2024Jan19;103(3):e36905
2. Menon G, Alkabban FM, Ferguson T. Breast Cancer. 2024 Feb 25. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing. 2024 Jan
3. Hong R, Xu B. Breast cancer: an up-to-date review and future perspectives. Cancer Commun (Lond). 2022Oct;42(10):913-936
4. Łukasiewicz S, Czeczelewski M, Forma A, Baj J, Sitarz R, Stanisławek A. Breast Cancer-Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies-An Updated Review. Cancers (Basel). 2021Aug25;13(17):4287
5. Mahdavi M, Nassiri M, Kooshyar MM, Vakili-Azghandi M, Avan A, Sandry R. et al. Hereditary breast cancer; Genetic penetrance and current status with BRCA. J Cell Physiol. 2019May;234(5):5741-5750
6. Zhao Y, Xu C. [Structure and function of angiotensin converting enzyme and its inhibitors]. Sheng Wu Gong Cheng Xue Bao. 2008Feb;24(2):171-6
7. Fountain JH, Kaur J, Lappin SL. Physiology, Renin Angiotensin System. 2023 Mar 12. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing. 2024 Jan
8. Fatehi R, Nouraei M, Panahiyan M, Rashedinia M, Firouzabadi N. Modulation of ACE2/Ang1-7/Mas and ACE/AngII/AT1 axes affects anticancer properties of sertraline in MCF-7 breast cancer cells. Biochem Biophys Rep. 2024May23;38:101738
9. Karnati S, Lüers G, Pfreimer S, Baumgart-Vogt E. Mammalian SOD2 is exclusively located in mitochondria and not present in peroxisomes. Histochem Cell Biol. 2013Aug;140(2):105-17
10. Huang M, Liu YU, Yao X, Qin D, Su H. Variability in SOD1-associated amyotrophic lateral sclerosis: geographic patterns, clinical heterogeneity, molecular alterations, and therapeutic implications. Transl Neurodegener. 2024May29;13(1):28
11. Crown AM, Roberts BL, Crosby K, Brown H, Ayers JI, Hart PJ, Borchelt DR. Experimental Mutations in Superoxide Dismutase 1 Provide Insight into Potential Mechanisms Involved in Aberrant Aggregation in Familial Amyotrophic Lateral Sclerosis. G3 (Bethesda). 2019Mar7;9(3):719-728
12. Archer SN, Schmidt C, Vandewalle G, Dijk DJ. Phenotyping of PER3 variants reveals widespread effects on circadian preference, sleep regulation, and health. Sleep Med Rev. 2018Aug;40:109-126
13. Wang Q, Liu H, Wang Z, Chen Y, Zhou S, Hu X. et al. Circadian gene Per3 promotes astroblastoma progression through the P53/BCL2/BAX signalling pathway. Gene. 2024Feb15;895:147978
14. Climent J, Perez-Losada J, Quigley DA, Kim IJ, Delrosario R, Jen KY. et al. Deletion of the PER3 gene on chromosome 1p36 in recurrent ER-positive breast cancer. J Clin Oncol. 2010Aug10;28(23):3770-8
15. Bièche I, Khodja A, Lidereau R. Deletion mapping of chromosomal region 1p32-pter in primary breast cancer. Genes Chromosomes Cancer. 1999;24(3):255-263
16. Pendergast JS, Niswender KD, Yamazaki S. Tissue-specific function of Period3 in circadian rhythmicity. PLoS One. 2012;7(1):e30254
17. Bae K, Jin X, Maywood ES, Hastings MH, Reppert SM, Weaver DR. Differential functions of mPer1, mPer2, and mPer3 in the SCN circadian clock. Neuron. 2001;30(2):525-536
18. Partch CL, Green CB, Takahashi JS. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014;24(2):90-99
19. Shih HC, Choo KB, Chang TJ, Yang MY, Shih MC, Yeh KT. et al. Disturbance of circadian gene expression in endometrial cancer: detection by real-time quantitative RT-PCR. Oncol Rep. 2005Dec;14(6):1533-8
20. Wang X, Yan D, Teng M, Fan J, Zhou C, Li D. et al. Reduced expression of PER3 is associated with incidence and development of colon cancer. Ann Surg Oncol. 2012Sep;19(9):3081-8
21. Wood PA, Yang X, Hrushesky WJ. Clock genes and cancer. Integr Cancer Ther. 2009Dec;8(4):303-8
22. Essobky H, Abdel-Megied A, El-Mezayen H, Farouk O, Refaat S, Hamed S. Is Angiotensin Converting Enzyme Insertion/Deletion (rs1799752) Polymorphism Associated with Breast Cancer Risk in Egyptian Population? Asian J. Biochem. Genet. Mol. Biol. 2022;10(3):8-22
23. Eskandari-Nasab E, Kharazi-Nejad E, Nakhaee A, Afzali M, Tabatabaei SP, Tirgar-Fakheri K. et al. 50-bp Ins/Del polymorphism of SOD1 is associated with increased risk of cardiovascular disease. Acta Med Iran. 2014;52(8):591-5
24. Fores-Martos J, Cervera-Vidal R, Sierra-Roca J, Lozano-Asencio C, Fedele V, Cornelissen S, Edvarsen H, Tadeo-Cervera I, Eroles P, Lluch A, Tabares-Seisdedos R, Falcó A. et al. Circadian PERformance in breast cancer: a germline and somatic genetic study of PER3VNTR polymorphisms and gene co-expression. NPJ Breast Cancer. 2021Sep10;7(1):118
25. Nyholt DR. A simple correction for multiple testing for single-nucleotide polymorphisms in linkage disequilibrium with each other. Am J Hum Genet. 2004;74(4):765-9
26. Li J, Ji L. Adjusting multiple testing in multilocus analyses using the eigenvalues of a correlation matrix. Heredity. 2005;95(3):221-7
27. Feng Y, Spezia M, Huang S, Yuan C, Zeng Z, Zhang L. et al. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018May12;5(2):77-106
28. Zuo X, Ren S, Zhang H, Tian J, Tian R, Han B. et al. Chemotherapy induces ACE2 expression in breast cancer via the ROS-AKT-HIF-1α signaling pathway: a potential prognostic marker for breast cancer patients receiving chemotherapy. J Transl Med. 2022Nov5;20(1):509
29. Rigat B, Hubert C, Alhenc-Gelas F, Cambien F, Corvol P, Soubrier F. An insertion/deletion polymorphism in the angiotensin I-converting enzyme gene accounting for half the variance of serum enzyme levels. J Clin Invest. 1990;86:1343-1346
30. Koh WP, Yuan JM, Sun CL, van den Berg D, Seow A, Lee HP, Yu MC. Angiotensin I-converting enzyme (ACE) gene polymorphism and breast cancer risk among Chinese women in Singapore. Cancer Res. 2003Feb1;63(3):573-8
31. Moghimi M, Kargar S, Jafari MA, Ahrar H, Jarahzadeh MH, Neamatzadeh H. et al. Angiotensin Converting Enzyme Insertion/Deletion Polymorphism is Associated with Breast Cancer Risk: A Meta-Analysis. Asian Pac J Cancer Prev. 2018Nov29;19(11):3225-3231
32. Singh A, Srivastava N, Amit S, Prasad SN, Misra MP, Ateeq B. Association of AGTR1 (A1166C) and ACE (I/D) Polymorphisms with Breast Cancer Risk in North Indian Population. Transl Oncol. 2018Apr;11(2):233-242
33. Yaren A, Turgut S, Kursunluoglu R, Oztop I, Turgut G, Kelten C. et al. Association between the polymorphism of the angiotensin-converting enzyme gene and tumor size of breast cancer in premenopausal patients. Tohoku J Exp Med. 2006Oct;210(2):109-16
34. van der Knaap R, Siemes C, Coebergh JW, van Duijn CM, Hofman A, Stricker BH. Renin-angiotensin system inhibitors, angiotensin I-converting enzyme gene insertion/deletion polymorphism, and cancer: the Rotterdam Study. Cancer. 2008Feb15;112(4):748-57
35. Li XL, Zheng ZJ, Qu HO. Lack of association of angiotensin-converting enzyme insertion/deletion polymorphism with breast cancer: An update meta-analysis based on 10405 subjects. J Renin Angiotensin Aldosterone Syst. 2015Dec;16(4):1095-100
36. Zhang Y, He J, Deng Y, Zhang J, Li X, Xiang Z. et al. The insertion/deletion (I/D) polymorphism in the Angiotensin-converting enzyme gene and cancer risk: a meta-analysis. BMC Med Genet. 2011Dec12;12:159
37. Ishfaq A, Ali E, Fatima W, Hussain N. Dr Genetic Association of Angiotensin Converting Enzyme I/D Gene Polymorphisms with Breast Cancer. Eur J Biol Biotechnol. 2020 1(4)
38. Papa L, Hahn M, Marsh EL, Evans BS, Germain D. SOD2 to SOD1 switch in breast cancer. J Biol Chem. 2014Feb28;289(9):5412-6
39. Gomez ML, Shah N, Kenny TC, Jenkins EC Jr, Germain D. SOD1 is essential for oncogene-driven mammary tumor formation but dispensable for normal development and proliferation. Oncogene. 2019Jul;38(29):5751-5765
40. Milani P, Gagliardi S, Bongioanni P, Grieco GS, Dezza M, Bianchi M. et al. Effect of the 50 bp deletion polymorphism in the SOD1 promoter on SOD1 mRNA levels in Italian ALS patients. J Neurol Sci. 2012;313(1-2):75-8
41. Saify K, Saadat M. Influence of a 50bp Ins/Del polymorphism at promoter of the superoxide dismutase-1 on gene expression and risk of heroin dependency. Environ Health Prev Med. 2017;22(1):4
42. Mahjoub G, Saadat I. Genetic Polymorphisms in CAT-21A/T and SOD1 50 bp I/D Genes with the Risk of Breast Cancer. Gene Reports. 2020;19:100609
43. Gallegos-Arreola MP, Ramírez-Hernández MA, Figuera LE, Zúñiga-González GM, Puebla-Pérez AM. The rs2234694 and 50 bp Insertion/Deletion polymorphisms of the SOD1 gene are associated with breast cancer risk in a Mexican population. Eur Rev Med Pharmacol Sci. 2020Aug;24(15):8017-8027
44. Liu Y, Wu Z, Li Y, Zhang J, Gao Y, Yuan G. et al. PER3 plays anticancer roles in the oncogenesis and progression of breast cancer via regulating MEK/ERK signaling pathway. J Chin Med Assoc. 2022Nov1;85(11):1051-1060
45. Blakeman V, Williams JL, Meng QJ, Streuli CH. Circadian clocks and breast cancer. Breast Cancer Res. 2016Sep2;18(1):89
46. Zhu Y, Brown HN, Zhang Y, Stevens RG, Zheng T. Period3 structural variation: a circadian biomarker associated with breast cancer in young women. Cancer Epidemiol Biomarkers Prev. 2005Jan;14(1):268-70
47. Wirth M, Burch J, Violanti J, Burchfiel C, Fekedulegn D, Andrew M. et al. Association of the Period3 clock gene length polymorphism with salivary cortisol secretion among police officers. Neuro Endocrinol Lett. 2013;34(1):27-37
48. Wirth MD, Burch JB, Hébert JR, Kowtal P, Mehrotra-Kapoor A, Steck SE. et al. Case-control study of breast cancer in India: Role of PERIOD3 clock gene length polymorphism and chronotype. Cancer Invest. 2014Aug;32(7):321-9
49. Barlattani T, Soltmann B, D'Amelio C, Socci V, Pacitti F, Pompili M. et al. The influence of PER3 VNTR genotypes on the age of onset in a group of bipolar I disorder patients: an exploratory study. Int J Bipolar Disord. 2024Jul11;12(1):25
50. Chen B, Cole JW, Grond-Ginsbach C. Departure from Hardy Weinberg Equilibrium and Genotyping Error. Front Genet. 2017Oct31;8:167
51. Abramovs N, Brass A, Tassabehji M. Hardy-Weinberg Equilibrium in the Large Scale Genomic Sequencing Era. Front Genet. 2020Mar13;11:210
52. Fardo DW, Becker KD, Bertram L, Tanzi RE, Lange C. Recovering unused information in genome-wide association studies: the benefit of analyzing SNPs out of Hardy-Weinberg equilibrium. Eur J Hum Genet. 2009;17(12):1676-82
53. Hosking L, Lumsden S, Lewis K, Yeo A, McCarthy L, Bansal A. et al. Detection of genotyping errors by Hardy-Weinberg equilibrium testing. Eur J Hum Genet. 2004;12(5):395-9
54. Wigginton JE, Cutler DJ, Abecasis GR. A note on exact tests of Hardy-Weinberg equilibrium. Am J Hum Genet. 2005;76(5):887-93
55. Zintzaras E. Impact of Hardy-Weinberg equilibrium deviation on allele-based risk effect of genetic association studies and meta-analysis. Eur J Epidemiol. 2010;25(8):553-60
Author contact
![]() Corresponding author: Prof. Laith Al-Eitan at Department of Biotechnology and Genetic Engineering, Jordan University of Science and Technology. P.O. Box 3030, Irbid 22110, Jordan. Tel: + (962)-2 -7201000. Fax: + (962)-2-7201071. Email: lneitanedu.jo.
Corresponding author: Prof. Laith Al-Eitan at Department of Biotechnology and Genetic Engineering, Jordan University of Science and Technology. P.O. Box 3030, Irbid 22110, Jordan. Tel: + (962)-2 -7201000. Fax: + (962)-2-7201071. Email: lneitanedu.jo.