Impact Factor ISSN: 1837-9664

 Global reach, higher impact
Global reach, higher impactJ Cancer 2015; 6(4):351-359. doi:10.7150/jca.11093 This issue Cite
Research Paper
Expression and Function of HAX-1 in Human Cutaneous Squamous Cell Carcinoma
Xingyu Li1*, Tiejun Li1,2*, Bo You3, Yin Shan3, Si Shi3, Xiaolei Cao1 ![]() , Li Qian4
, Li Qian4 ![]()
1. Department of Pathological Anatomy, Medical School of Nantong University, Nantong 226001, Jiangsu, China;
2. Small RNA Technology and Application Institute, Nantong University, E&T Development Area, Nantong 226016, Jiangsu, China;
3. Medical School of Nantong University, Nantong 226001, Jiangsu, China;
4. Department of Oncology, Affiliated Hospital of Nantong University, Nantong 226001, Jiangsu, China.
*The first two authors have contributed equally to this work.
Received 2014-11-18; Accepted 2015-1-1; Published 2015-2-20
Abstract
HAX-1 is an anti-apoptotic factor and overexpressed in many types of cancers. However, the functional role of HAX-1 in human cutaneous squamous cell carcinoma (cSCC) remains unclear. Our aim was to investigate the expression of HAX-1 in cSCC and its relationship with the development of cSCC. HAX-1 expression in cSCC tissues and in-vitro cell models were evaluated by real-time quantitative PCR (RT-qPCR), Western blot and immunohistochemistry. And the RNAi strategy was used to observe the relationship of HAX-1 and cSCC in A431 cells. The mRNA and protein level of HAX-1 were significantly higher in cSCC compared with normal tissue. There were significant differences in thickness (P=0.014), differentiation (P=0.027) and TNM stages (P=0.007). After knockdown the expression of HAX-1 by siRNA, the proliferation and the motility of A431 cell was inhibited obviously, and the apoptosis of A431 cells were induced too. HAX-1 may be a risk factor for patients with cSCC. As a potential tumor promoter in cSCC, HAX-1 may be a novel potential therapeutic target for cSCC treatment and deserves further investigation.
Keywords: HAX-1, Cutaneous squamous cell carcinoma (cSCC), RNA interference (RNAi), A431.
Introduction
Skin cancer, which can be categorized as melanoma and non-melanoma skin carcinoma, is one of the most common malignancies in the world [1]. Non-melanoma includes basal and squamous cell carcinoma. Because of the changing of lifestyle, cutaneous squamous cell carcinoma (cSCC) leads to a high morbidity rate [2]. The major risk factors of cSCC include endogenous genetic factors and exogenous factors such as sun exposure, chemical carcinogens, ionizing radiation and other environmental stress [3]. The incidence of cSCC is increasing year by year, although the cure rate can be high with surgical treatment and radiotherapy, some patients still develop metastasis and the quality of life may be influenced [4]. Therefore, better understanding of the pathogens and the molecular mechanism is important for the early diagnosis and treatment of cSCC.
HS1 associated protein X-1 (HAX-1), a novel intracellular protein was first found as a partner of hematopoietic cell signaling protein HS1, and it mainly locates in the mitochondrial and is a kind of multifunctional protein [5]. HAX-1 is regarded as an important target gene of cancer because it is involved in the regulation of apoptosis, cell migration and metastasis [6, 7]. Several studies also have proved that HAX-1 was over-expressed in a variety of tumors, including colorectal cancer [8], esophageal squamous carcinoma (ESCC) [9], oral squamous cell carcinoma [7], hepatoma [10], breast cancer [11], lung cancer [11], B lymphoma [12], melanoma [13], and etc. Currently, Hax-1 was found to suppress melanoma cell apoptosis and promote its growth [13], however, the expression pattern and possible roles of HAX-1 in human cSCC have not been investigated.
In the present study, we evaluated the expression level of HAX-1 in cSCC cell lines and clinical tissues. Then we assessed clinical and pathological roles of HAX-1 in cSCC. Furthermore, the biological functions of HAX-1 expression on cell proliferation, apoptosis, adhesion and motility abilities in vitro, suggested the possibility of HAX-1 as a potential target in cSCC therapy.
Materials and Methods
Clinical Specimens and Immunohistochemical Staining
A total of 60 cases of cSCC tissues and 30 cases of normal skin tissues were obtained from the dermatological department of Nantong University Affiliated Hospital. Both tumor and normal skin tissues were histologically confirmed. All of patients were staged using the seventh edition of the International Union Against Cancer tumor-node-metastasis (TNM) staging system. Informed consent was obtained from each patient, and this study was approved by the Institute Research Ethics Committee of the Nantong University Nantong University Affiliated Hospital, China.
All resected samples were fixed with formalin and embedded in paraffin, then sectioned at 4-μm thickness. Envision Plus/Horseradish Peroxidase system (DAKO Corp, USA) was employed for immunohistochemical staining. The slides were incubated with anti-HAX-1 mouse polyclonal antibody (Santa Cruz, USA, dilution 1:200) overnight at 4 °C. After washing with Tris-buffered saline, the slides was incubated using the Envision Plus secondary antibody for 30 min, followed by diaminobenzidine for 5 min. Appropriate positive and negative controls (incubation with secondary antibody only) were stained in parallel.
The immunohistochemistry scoring results were evaluated independently by the pathologists without knowing the patient's clinic pathological outcomes, and then the results analyzed according to the intensity of the staining and the relative abundance of positive cells. The intensity was graded as 0 (negative), 1 (weakly positive), 2 (moderately positive) or 3 (strongly positive). The percentage of positive cells was graded as 0 (≤5%), 1 (6-25%), 2 (26-50%), 3 (51-75%), 4 (>75%). The scoring results were obtained by intensity and percentage score. The expression level of HAX-1 was defined as follows: “-” (negative, score of 0), “+” (weakly positive, score of 1-4), “++” (positive, score of 5-8), “+++” (strongly positive, score of 9-12).
Cell Culture and siRNA Transfection
Human cSCC cell line A431 and human keratinocyte cell line HaCaT (cells were all obtained from Insitute of Biochemistry and Cell Biology of the Chinese Academy of Sciences, Shanghai, China) were cultured in Dulbecco's modified Eagle's medium (DMEM) (Gibco, USA) with 10% fetal bovine serum (FBS) (Gibco, USA) at 37 °C in 5% CO2 incubator.
The cells were transfected with siRNAs using Lipofectamine® 2000 transfection reagent (Life Technologies, USA) according to the manufactures' instructions. Four siRNAs targeting HAX-1 (Accession No. NM_006118 form NCBI GenBank) were designed and the BLAST analysis of siRNA sequences was carried out to ensure on homology with others genes in human. And the negative control siRNA (NC_siR) was also designed. The sequences of siRNAs were show in Table 1. All siRNAs were designed and synthesized by Biomics Biotechnologies Co., Ltd (China).
Sequences of siRNAs were designed targeting HAX-1.
| siRNAs | Sequences (5'-3') |
|---|---|
| HAX_siR1 | Sense: GGAUACGUUUCCACGAUAAdTdT |
| Antisense: UUAUCGUGGAAACGUAUCCdTdT | |
| HAX_siR2 | Sense: GAGUGAUGCAAGAAGUGAAdTdT |
| Antisense: UUCACUUCUUGCAUCACUCdTdT | |
| HAX_siR3 | Sense: GAAGCAGAUAGCAGUCCUAdTdT |
| Antisense: UAGGACUGCUAUCUGCUUCdTdT | |
| HAX_siR4 | Sense: GGACUGUGGUGGACAGUGAdTdT |
| Antisense: UCACUGUCCACCACAGUCCdTdT | |
| NC_siR | Sense: UUCUCCGAACGUGUCACGUdTdT |
| Antisense: ACGUGACACGUUCGGAGAAdTdT |
Expression of HAX-1 in 60 cutaneous squamous carcinoma and 30 normal skin tissues.
| HAX-1 | P | ||||
|---|---|---|---|---|---|
| - | + | ++ | +++ | ||
| cSCC | 0 | 23 | 27 | 10 | <0.05* |
| Normal skin tissues | 27 | 2 | 1 | 0 | |
* represents statistical significance, P<0.05.
RT-qPCR
Total RNA of HaCaT and A431 cells were extracted using a TRIzol® reagent (Life Technologies, USA) according to the manufacturer's procedure. cDNA was produced using a Transcriptor First Strand cDNA Synthesis Kit (Roche, Germany). Then 480 SYBR Green I Master (Roche, Germany) was used for one-step RT-qPCR. The mRNA of HAX-1 was detected by forward primer, 5'-TCAATAGCATCTTCAGCGATATG-3' and reverse primer, 5'-GTCCCTCCCGTAGTCTCTC-3'. The thermal cycling profile consisted of 10 min at 94 °C for pre-denature, followed by 40 cycles of denature at 94 °C for 20 s, annealing at 58 °C for 20 s and extension at 72 °C for 30 s. All reactions were run in triplicate. Human glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (forward: 5'-GAAGGTGAAGGTCGGAGTC-3'; reverse: 5'-GAAGATGGTGATGGGATTTC-3') served as the internal control for mRNA determination of HAX-1. The experiment was performed in triplicate. The results were analyzed by 2-ΔΔCt method [14].
Western Blot
The transfected cells were harvested and lysed in ice-cold cell RIPA buffer (Pierce, USA) and then centrifuged at 10,000 rpm for 20 min at 4 °C. And the protein were separated by SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and electro-transferred to polyvinylidine difluoride filter (PVDF) membranes (Millipore, USA). The membranes were incubated with goat-anti-human HAX-1 (1:500 dilutions, Abcam, USA) and mouse-anti-human β-actin (1:200 dilution, Santa Cruz, USA) as internal control. After washing with TBST, the membrane was incubated with the secondary antibody conjugated to horseradish peroxidase (HRP) (donkey anti-goat IgG-HRP with 1:1,000 dilution for HAX-1; goat anti-mouse IgG-HRP with 1:2,000 dilution for β-actin) for about 1.5 h at room temperature, and then three times washed in TBST for 5 min, the specific proteins were detected with ECL plus kit (ZSbio, China).
Cell Viability Assay
Cell viability was measured using the Cell Counting Kit-8 (CCK-8) kit (Beyotime, China). A431 cells were seeded in triplicate at a concentration of 5×103 cells per well on a 96-well plate before transfection and growth to 70-80% confluence for 24 h. After treatment with siRNAs for 0, 24, 48, 72 and 96 h, 10 μL of CCK-8 reagent was added to each well containing 100 μL culture medium and cells were further incubated for 2 h at 37 °C. Then the optical density (OD) was measured at 450 nm using a Fluorescence Spectrophotometer (HITACHI, Japan).
Scratch Wound Healing Assay
A431 cells were plated and transfected with siRNAs as described above in a 12-well plate at the density of 3×105cells per well. After 48 h, wound was made through confluent monolayer cells with a pipette tip and cells were washed with PBS and then DMEM medium with no FBS. Photographs of cells were taken at 0, 24, 48, and 72 h to monitor cell movements.
Matrigel Invasion Assay
Cell invasion assay was performed using a 24 well transwell chamber (8-μm pore size, Corning, USA), and the upper chambers were coated with 50 μL of 0.5 mg/ml Matrigel (BD Biosciences, USA). After transfection with siRNAs for 48 h, A431 cells were treated with 0.25% trypsin and added to the upper chamber in serum-free DMEM medium at a concentration of 1×106 cells per mL. DMEM medium with 10% FBS was added to the lower chamber as the chemo attractant. The cells on the top surface of the membrane were carefully removed at 24 h after transfection with a cotton tip. Cells on the transwell chambers were fixed by 10% formaldehyde and stained by 0.5% crystal violet. Invasion cells on the bottom surface of the membrane were counted in five random fields (100×) per membrane, and the average number of migrating cells per field was calculated.
Cell Apoptosis Assay
Apoptosis of the A431 cells treated by siRNAs was determined using Annexin V-FITC/PI double staining and flow cytometry (FCM) analysis. Briefly, 1×106 cells per well after 48 h with different treatments described above were harvested and washed with PBS in 6 well plates. The cells were washed with PBS and resuspended in Binding Buffer, followed incubated with Annexin V-FITC conjugate and PI for 15 min at room temperature. Cells were analyzed by FCM analysis using BD CELLQuest software (BD Biosciences, USA).
Caspase-3 Activity Assay
Caspase-3 enzymatic activity was determined using an Active Caspase 3 Human ELISA Kit (Abcam, USA) according to the manufacturer's protocol. Briefly, A431 cells were treated with siRNAs as described above. After different treatments, cell lysates were prepared and Detector Antibody was added to cell lysates. The mixtures were incubated at room temperature for 1 h and then the HRP Label was added for 1 h at room temperature. Following, the samples were measured with an ELISA reader (Bio-Tek, USA) at an absorbance of 405 nm for caspase-3 activity.
Statistical Analysis
All experiments were performed independently three times, the results were shown as mean values±standard deviation (SD), and statistical analyses were performed using SPSS19.0 software. Results were analyzed using chi-square test and Student's t-test to assess statistical significance. All P values are based on a two-sided statistical analysis and P<0.05 was considered to indicate statistical significance.
Results
HAX-1 was overexpressed in cSCC cell
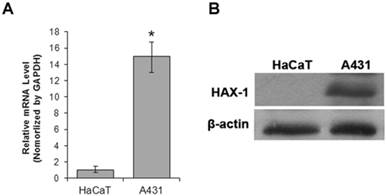
The expression level of HAX-1 in cSCC cell line A431 was detected compared with human keratinocyte cell line HaCaT using Western blot and RT-qPCR methods. The results of RT-qPCR and Western blot assay showed that the mRNA and protein expression level of HAX-1 was effectively higher in A431cells (Fig. 1), compared with HaCaT cells.
HAX-1 was up-regulated in cSCC. (A) RT-qPCR was used to detect the mRNA expression of HAX-1. *P<0.05 compared with expression level in HaCaT cells, (B) Western blotting analysis of Hax-1 expression in A431 cell lines and HaCaT cell lines.
Correlation of the expression of HAX-1 with clinico pathological factors in cSCC
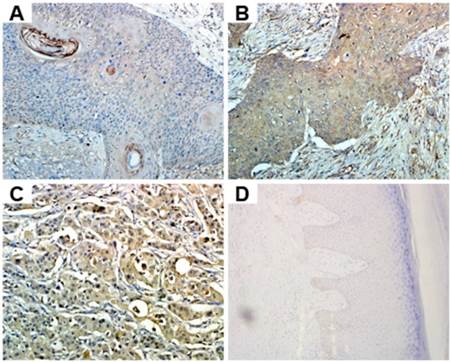
Because of the differential expression of HAX-1 between cancer cells and normal cells, we hypothesized it might play an important role in cSCC. Overexpression of HAX-1 in cSCC tissues was further tested by immunohistochemistry (Fig. 2). We found that the moderate or strong staining of HAX-1 in cSCC tissues was 61.67 % (37/60), while normal was 3.33% (1/30). Thus, the expression of HAX-1 was higher in cSCC tissues than the normal skin tissues (P<0.05). According to previous studies, we demonstrated that HAX-1 was over-expressed in cSCC tissues.
The expression of HAX-1 was analyzed by immunohistochemistry. (A) well-differentiated cSCC, (B) moderately differentiated cSCC, (C) poorly differentiated cSCC, (D) normal skin. (200×).
The level of HAX-1 expression was associated with the clinicopathological factors of cSCC. The clinical and pathological data of 60 cSCC patients was displayed in Table 3. There was no statistical difference found between HAX-1 positive expression and the characteristics of gender, age, sunlight exposure. However, HAX-1 overexpression in cSCC was strongly associated with thickness (P=0.014), differentiation (P=0.027) and TNM stages, (P=0.007).
The association between HAX-1 expression and clinical pathological factors in cSCC.
| Clinicopathological parameters | No. Case | HAX-1 Expression, n | P | ||||
|---|---|---|---|---|---|---|---|
| -~+ | ++ | +++ | |||||
| Gender | 0.767 | ||||||
| Male | 33 | 14 | 14 | 5 | |||
| Female | 27 | 9 | 13 | 5 | |||
| Age | 0.553 | ||||||
| <60 | 5 | 2 | 3 | 0 | |||
| ≥60 | 55 | 21 | 24 | 10 | |||
| Thickness | 0.014* | ||||||
| <2 mm | 14 | 9 | 5 | 0 | |||
| 2-5 mm | 15 | 2 | 11 | 2 | |||
| >5 mm | 31 | 12 | 11 | 8 | |||
| TNM stages | 0.027* | ||||||
| Tis | 6 | 6 | 0 | 0 | |||
| I | 30 | 12 | 14 | 4 | |||
| II | 20 | 5 | 10 | 5 | |||
| III | 4 | 0 | 3 | 1 | |||
| Differentiation | 0.007* | ||||||
| Well | 25 | 12 | 12 | 1 | |||
| Moderately | 25 | 11 | 10 | 4 | |||
| Poorly | 10 | 0 | 5 | 5 | |||
| Sites | 0.807 | ||||||
| Sunlight exposure | 58 | 22 | 26 | 10 | |||
| Non exposure | 2 | 1 | 1 | 0 | |||
* represents statistical significance, P<0.05.
Inhibition effects of pre-designed siRNAs targeting HAX-1
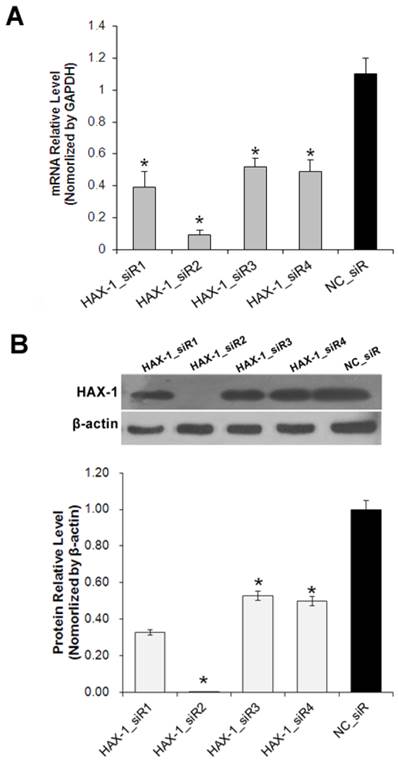
To investigate the role of HAX-1 in cSCC, the expression of HAX-1 was knocked down using RNAi method. A431 cells were transfected with four pre-designed siRNAs. As shown in Fig. 3, the mRNA (Fig. 3A) and protein (Fig. 3B) level of HAX-1 were both inhibited significantly by HAX-1_siR2.
Expression of HAX-1 in A431 cells is effectively down-regulated by specific HAX-1 siRNAs. (A) HAX-1 knockdown efficiency at mRNA level was detected by RT-qPCR, (B) HAX-1 knockdown efficiency at protein level was detected by Western blot. *P<0.05 compared with NC_siR.
HAX-1 promotes the proliferation of cSCC cells
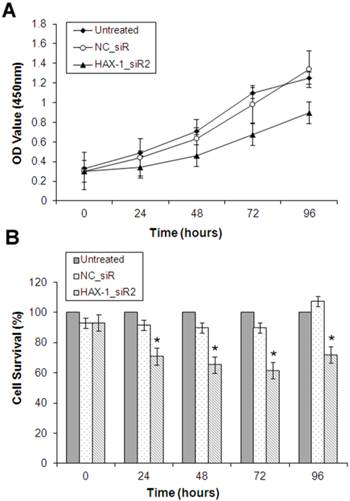
Cell Counting Kit 8 (CCK-8) assay was used to examine the role of HAX-1 on proliferation of A431 cells with the transfection of siRNAs. The cell proliferation was significantly decreased after treatment with HAX-1_siR2 at 24, 48, 72 and 96 h compared with the transfection of NC_siR and untreated groups (Fig. 4).
The effect of HAX-1 on cell proliferation analyzed by CCK8 assay. (A) Growth curve of A431 cells was shown for each treatment at 0, 24, 48, 72, and 96 h. (B) The inhibition rate of A431 cells with different treatments. *P<0.05 compared with NC_siR and untreated group.
HAX-1 promotes the motility of cSCC cells
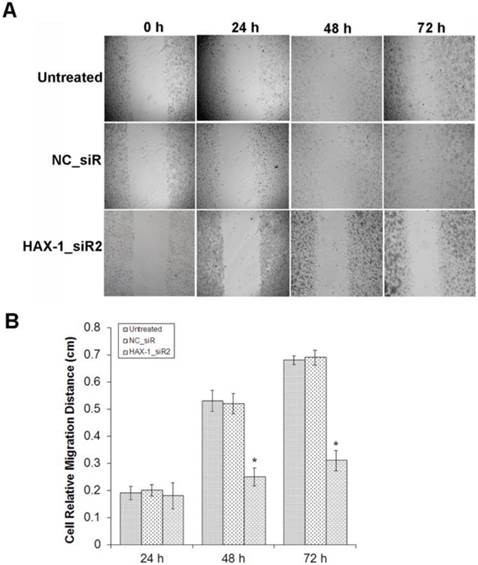
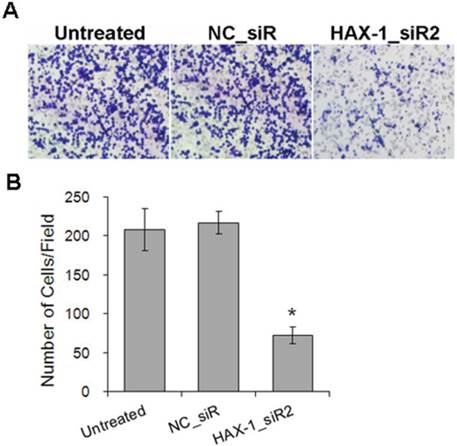
Moreover, to determine whether HAX-1 plays a crucial role in cell motility using the scratch wound healing and matrigel invasion assay. Scratch wound healing assay was used to evaluate the migration ability of A431 cells with different treatments. We observed the scratch caused in groups of untreated and NC_siR nearly closed completely after 72 h, but the cells in treatment with HAX-1_siR2 was not able to move toward the center of the wound (Fig. 5A). HAX-1_siR2 treated cells showed a decrease in wound healing ability compared to NC_siR treated and untreated group (P<0.05) (Fig. 5B). The result of transwell assay showed that the migration of the cells with HAX-1_siR2 was significantly affected at 24 h post-transfection (P<0.05) (Fig. 6). Moreover, there was no significant difference in HAX-1 expression and migration between NC_siR and untreated cells (P>0.05).
The migration inhibition effects of A431 cells by HAX-1_siR2 was determined. (A) The cells with different treatments at 0, 24, 48, and 72 h (40×), (B) Cell relative migration distances in different time point treatments were presented as mean±SD. *P<0.05 compared with NC_siR and untreated group.
A431 cells invasion inhibited by HAX-1_siR2 was determined using Transwell assay. (A) Representative field photograph of transwell assay of A431 cells under different treatments after 72 h (100×), (B) The numbers of invaded cells were calculated by averaging the total number of cells from 5 fields. *P<0.05 compared with NC_siR and untreated group.
HAX-1 plays a key role in the apoptosis of cSCC cells
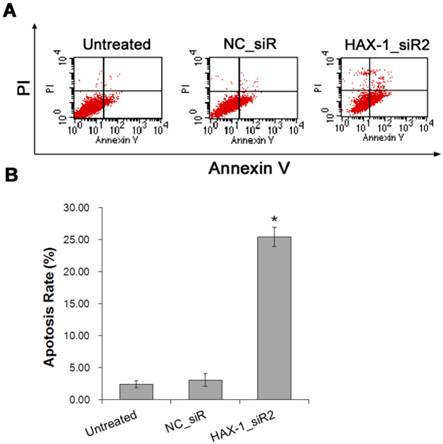
Furthermore, the role of HAX-1 on A431 cells apoptosis were examined by Annexin V-FITC/PI double staining and FCM analysis to evaluate the ability of HAX-1_siR2 on inducing A431 cells apoptosis. As shown in Fig. 7, treatment with siRNA resulted in a significant increase of apoptosis compared with that of untreated cells (P<0.05).
A431 cells apoptosis inhibited by HAX-1_siR2 was detected by Annexin V-FITC/PI double staining and FCM analysis. (A) Apoptosis rates were measured by FCM analysis after AnnexinV/PI staining, (B) Percentage of dual-positive (Annexin V and PI were positive) cells from three independent experiments was quantified and presented as mean±SD. *P<0.05 compared with NC_siR and untreated group.
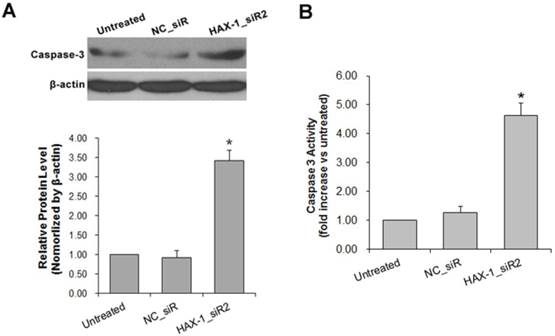
The expression of caspase-3 also detected in A431 cells with HAX-1 siRNA treated. Caspase-3 is an apoptosis marker which was up-regulated at the end stage of apoptosis cascades. The result of Western blot showed that the expression of active caspase-3 was increased in the HAX-1_siR2 treated group compared with untreated and NC_siR treated group (Fig. 8A). The enzymatic activity of caspase-3 was measured meanwhile, and the result showed that the enzymatic activity of caspase-3 was increased significantly in the HAX-1_siR2 treated group compared with untreated and NC_siR treated group (Fig. 8B).
Detection of Caspase-3 activation. (A) The expression of Caspase-3 in A431 cells with different treatments were detected by Western blot, (B) The expression of Caspase-3 in A431 cells with different treatments were detected by ELISA assay. *P<0.05 compared with NC_siR and untreated group.
Discussion
Cutaneous squamous cell carcinoma (cSCC) is one of the most common cancers in the world, and it is associated with a substantial risk of metastasis, the prognosis is poor for patients with tumors with regional or distant spread [2], so early diagnosis and treatment may be important for the patients.
HAX-1, as a multifunctional protein, is associated with many cellular proteins and viral proteins. Many studies showed that HAX-1 was involved in multiple signaling pathways and cellular processes including cancer cell apoptosis and migration. HAX-1 is over-expressed in a variety of tumors. In colorectal cancer, the mRNA expression level of HAX-1 was higher in tumor tissues than matched noncancerous tissues; its over-expression was significantly associated with lymph node metastasis and TNM stages [8]. In ESCC, high expression of HAX-1 was associated with lymph node metastasis and survival in the patients with ESCC [9]. In this study, we characterized HAX-1 expression in cSCC. We first chose A431 cell as the most suitable model of cSCC for experiments. The expression level of HAX-1 protein and mRNA were significantly higher in A431 cell line than normal cell line HaCaT. The overexpression of HAX-1 in cSCC was also confirmed at the protein level by immunohistochemistry. The HAX-1 expression was clearly up-regulated in most tumor tissue sections and the immuno-reactivity mainly in cytoplasm of the cells, which is consistent with previous findings that HAX-1 associating with cytoplasmic structures [15-17]. Besides that, we also found that there were significant differences in thickness, differentiation and TNM stages between high HAX-1 expression group and low HAX-1 expression group.
As a protein partner of HS1, HAX-1 also binds the Cortactin, which promotes tumor invasiveness and metastasis [18]. HAX-1 was also be demonstrated to play a role in regulation of carcinoma cell migration and invasion via clathrin-mediated endocytosis of integrin alphavbeta6 [7] and regulate human endothelial cell migration in vitro by interacting with the IL-1a precursor [19]. Sun et al. [9] have shown that HAX-1 promoted the proliferation, chemoresistance, invasion, and tumorigenicity of ESCC cells. In melanoma, HAX-1 has also been found to suppress melanoma cell apoptosis and promote its growth [13]. These studies suggest that HAX-1 plays an important role in tumorigenesis and tumor metastasis. Thus, to further investigate the role of HAX-1 in cSCC, we determined the effect of HAX-1 on behavior of cSCC cells. HAX-1 was down-regulated via RNAi strategy, the siRNAs that targeting HAX-1 were designed and screened in cSCC cells, and the best silencing siRNA (HAX-1_siR2) was used for following observation. After HAX-1 was silenced by HAX-1_siR2, the proliferation, migration, invasion of A431 cells were all inhibited, and the apoptosis of A431 cells were induced also. All the results suggest that the role of HAX-1 might as a potential tumor promoter in cSCC.
In summary, our findings indicate that HAX-1 may be a crucial factor in the development of cSCC and contributes to the cell proliferation, apoptosis and motility in cSCC cells. Therefore, HAX-1 might be a potential target for cSCC therapy using an RNAi-based approach alone or in combination with chemotherapeutics. Further investigations also should be done in vivo to make clear the mechanism of HAX-1 in initiation and progression of cSCC.
Acknowledgements
This work was supported by grants from Chinese National Natural Science Foundation (Nos. 81172841, 81202368 and 81471603), China Postdoctoral Science Foundation (No. 2013M541708); the project of “333 Natural Science Foundation” of Jiangsu Grant (No. BRA2013286); the ''Top Six Types of Talents'' Financial Assistance of Jiangsu Province Grant (No. 6); the project of Jiangsu Provincial Health Department (No. Z201005); the innovative project of Nantong University postgraduate students (No. 13025043); and the Jiangsu province's Outstanding Medical Academic Leader Program (No. LJ201136).
Competing Interests
The authors have declared that no competing interest exists.
References
1. Hao ZF, Ao JH, Zhang J. et al. ATF3 Activates Stat3 Phosphorylation through Inhibition of p53 Expression in Skin Cancer Cells. Asian Pac J Cancer Prev. 2013;14:7439-44
2. Alam M, Ratner D. Cutaneous squamous-cell carcinoma. N Engl J Med. 2011;344:975-83
3. Gordon R. Skin cancer: an overview of epidemiology and risk factors. Semin Oncol Nurs. 2013;29:160-9
4. Ratushny V, Gober MD, Hick R. et al. From keratinocyte to cancer: the pathogenesis and modeling of cutaneous squamous cell carcinoma. J Clin Invest. 2012;122:464-72
5. Suzuki Y, Demoliere C, Kitamura D. et al. HAX-1, a novel intracellular protein, localized on mitochondria, directly associates with HS1, a substrate of Src family tyrosine kinases. J Immunol. 1997;158:2736-44
6. Shaw J, Kirshenbaum LA. HAX-1 represses postmitochondrial caspase-9 activation and cell death during hypoxia-reoxygenation. Circ Res. 2006;99:336-8
7. Ramsay AG, Keppler MD, Jazayeri M. et al. HS1-associated protein X-1 regulates carcinoma cell migration and invasion via clathrin-mediated endocytosis of integrin alphavbeta6. Cancer Res. 2007;67:5275-84
8. Wei XJ, Li SY, Yu B. et al. Expression of HAX-1 in human colorectal cancer and its clinical significance. Tumour Biol. 2014;35:1411-5
9. Sun SJ, Feng L, Zhao GQ. et al. HAX-1 promotes the chemoresistance, invasion, and tumorigenicity of esophageal squamous carcinoma cells. Dig Dis Sci. 2012;57:1838-46
10. Banerjee A, Saito K, Meyer K. et al. Hepatitis C virus core protein and cellular protein HAX-1 promote 5-fluorouracil-mediated hepatocyte growth inhibition. J Virol. 2009;83:9663-71
11. Trebinska A, Rembiszewska A, Ciosek K. et al. HAX-1 overexpression, splicing and cellular localization in tumors. BMC cancer. 2010;10:76
12. Kwiecinska A, Ottosson-Wadlund A, Ceder R. et al. HAX-1 expression in human B lymphoma. Leukemia. 2011;25:868-72
13. Li WB, Feng J, Geng SM. et al. Induction of apoptosis by Hax-1 siRNA in melanoma cells. Cell Biol Int. 2009;33:548-54
14. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402-8
15. Simmen T. Hax-1: a regulator of calcium signaling and apoptosis progression with multiple roles in human disease. Expert Opin Ther Targets. 2011;15:741-51
16. Yedavalli VS, Shih HM, Chiang YP. et al. Human immunodeficiency virus type 1 Vpr interacts with antiapoptotic mitochondrial protein HAX-1. J Virol. 2005;79:13735-46
17. Grzybowska EA, Zayat V, Konopiński R. et al. HAX-1 is a nucleocytoplasmic shuttling protein with a possible role in mRNA processing. FEBS J. 2013;280:256-72
18. Gomathinayagam R, Muralidharan J, Ha JH. et al. Hax-1 is required for Rac1-Cortactin interaction and ovarian carcinoma cell migration. Genes Cancer. 2014;5:84-99
19. McMahon GA, Garfinkel S, Prudovsky I. et al. Intracellular precursor interleukin (IL)-1alpha, but not mature IL-1alpha, is able to regulate human endothelial cell migration in vitro. J Biol Chem. 1997;272:28202-5
Author contact
![]() Corresponding authors: Xiaolei Cao, Ph.D. Department of Pathological Anatomy, Medical School of Nantong University, 19 Qixiu Road, Nantong 226001, Jiangsu, China. Email: xiaoleiedu.cn; Li Qian, Ph.D. Department of Oncology, Affiliated Hospital of Nantong University, Nantong 226001, Jiangsu, China. Email: ntqianlicom; Tel: +86-513-85051729; Fax: +86-513-85051729.
Corresponding authors: Xiaolei Cao, Ph.D. Department of Pathological Anatomy, Medical School of Nantong University, 19 Qixiu Road, Nantong 226001, Jiangsu, China. Email: xiaoleiedu.cn; Li Qian, Ph.D. Department of Oncology, Affiliated Hospital of Nantong University, Nantong 226001, Jiangsu, China. Email: ntqianlicom; Tel: +86-513-85051729; Fax: +86-513-85051729.